对抗知识焦虑,从看懂这条开始
App 下载
做了10年RNA剪刀的它,竟管起了脂肪合成
COPII囊泡系统|SREBP1|脂肪生成调控|Drosha蛋白|分子细胞生物学|生命科学
对抗知识焦虑,从看懂这条开始
App 下载COPII囊泡系统|SREBP1|脂肪生成调控|Drosha蛋白|分子细胞生物学|生命科学
当你大快朵颐高脂高糖的蛋糕时,肝脏里的一场分子狂欢正在失控——脂肪像吹气球一样在肝细胞里堆积,胰岛素的指令越来越难传递,脂肪肝和代谢紊乱的阴影悄然逼近。过去我们以为,这场狂欢的核心是脂肪调控因子SREBP1,以及负责转运它的COPII囊泡系统。但空军军医大学杨倩团队的新研究,却揪出了一个所有人都没想到的「幕后推手」:那个在细胞核里剪了10年RNA的Drosha,居然跑到细胞质里,靠「握手」而非「剪刀」,直接操控了脂肪生成的开关。它到底是怎么跨界的?
Drosha,这个名字在分子生物学界一直和「miRNA合成」绑定——它是细胞核里的III型核糖核酸酶,和搭档DGCR8组成「微处理器」,像精准剪刀一样把长长的初级miRNA剪成短小的前体,启动整个miRNA调控网络。这是它干了十几年的「本职工作」,没人怀疑过它的身份。
但这次研究团队用RNA测序对比了Drosha和Dicer(miRNA合成的另一个关键酶)缺失的细胞,发现了一个奇怪的差异:Drosha缺失的细胞里,脂肪合成相关基因的变化幅度远大于Dicer缺失的细胞。这说明,Drosha在miRNA之外,还干着别的活。
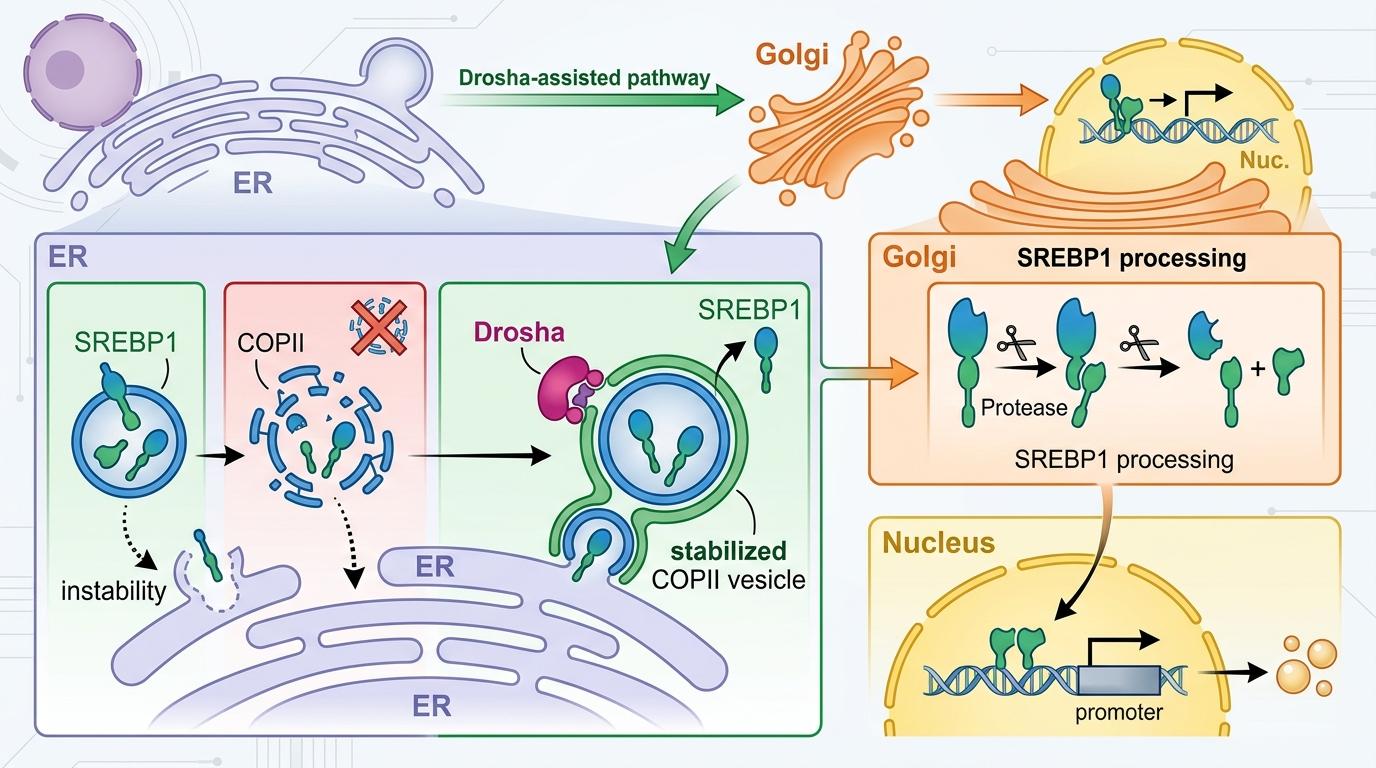
进一步的实验更颠覆认知:在细胞质里,Drosha完全丢掉了「剪刀」的身份——它不需要切割任何RNA,只靠自己N端那个富含精氨酸/丝氨酸的RS结构域,就能和COPII囊泡的外层亚基Sec31A紧紧「握手」。这个握手直接改变了COPII囊泡的命运:原本Sec31A会加速囊泡的解离,而Drosha的结合却抑制了这个过程,让囊泡能稳稳地带着SREBP1从内质网运到高尔基体。
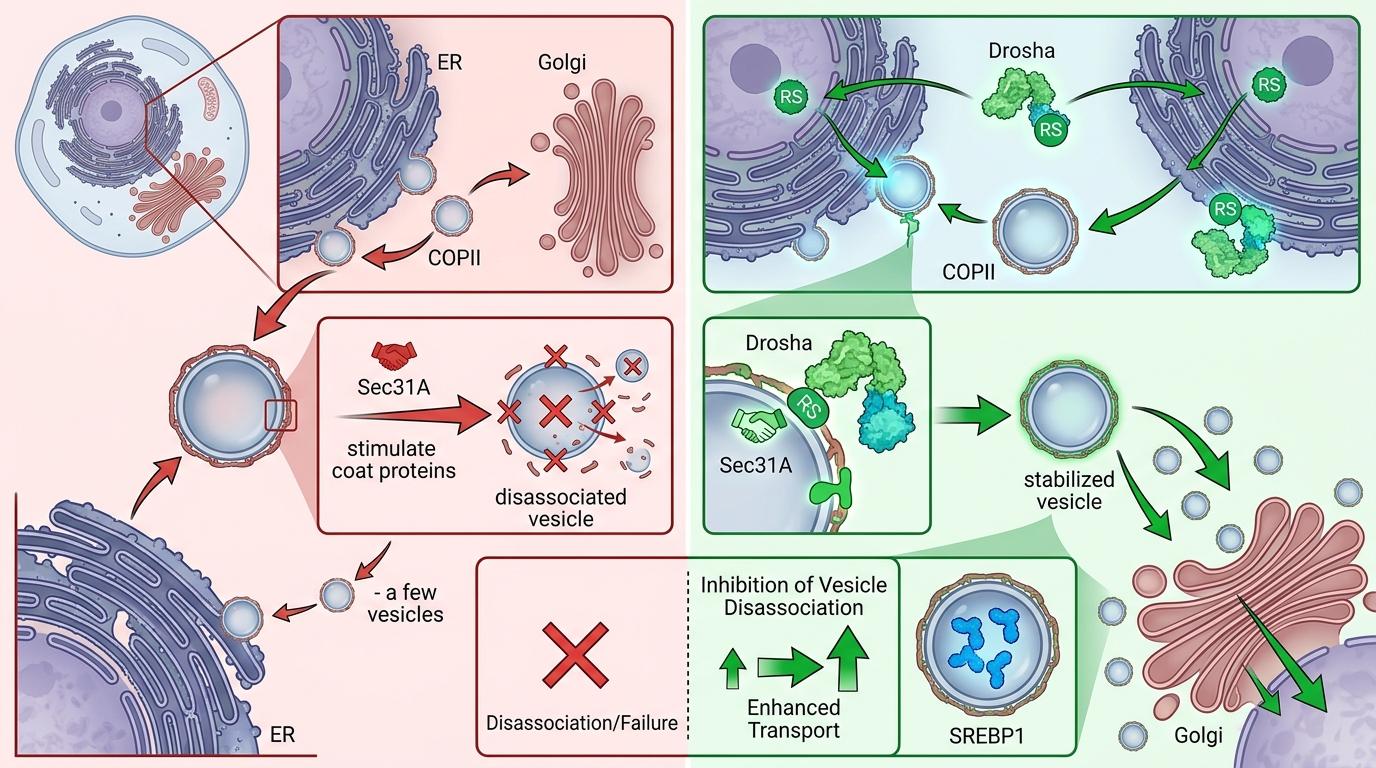
你可以把COPII囊泡想象成快递盒,SREBP1是要送进细胞核的「脂肪生成指令」。原本这个快递盒刚装好就容易散架,Drosha的出现就像给盒子缠了一圈胶带,让它能完整送到高尔基体这个「中转站」,在这里SREBP1被剪切成活性形式,直奔细胞核下令合成脂肪。
Drosha的跨界工作,还得靠胰岛素来发号施令。当我们吃完高糖食物,胰岛素会激活细胞里的Akt激酶,这个激酶像个精准的「磷酸化工匠」,会给Drosha RS结构域上的Ser237位点打上一个磷酸化标记。
这个标记至关重要:它就像给Drosha的RS结构域装了个「磁吸」,让它和Sec31A的结合力瞬间增强,快递盒的胶带缠得更紧,SREBP1的转运效率大幅提升,脂肪合成的油门被一脚踩到底。在高脂高糖饮食的小鼠体内,这条Akt-Drosha-SREBP1轴被过度激活,肝脏里的脂肪像洪水一样堆积,胰岛素也渐渐失去了作用。
研究团队做了两个关键实验:一是把小鼠肝脏里的Drosha敲除,二是给小鼠注射能阻断Akt给Drosha打标记的干扰肽。结果都指向同一个结论:这条轴一旦被抑制,小鼠肝脏的脂肪沉积明显减少,胰岛素抵抗也得到了改善。这意味着,Drosha不再只是个RNA加工酶,它成了连接胰岛素信号和脂肪生成的关键节点。
不过,这个机制也有它的局限:目前的实验只在肝脏和小鼠模型中得到验证,Drosha的跨界功能在脂肪组织、肌肉等其他代谢器官里是否同样存在,还需要更多研究。而且,干扰肽的临床转化还面临着递送效率、特异性等诸多问题,距离真正的药物还有一段路要走。
这次的发现,只是掀开了Drosha非经典功能的冰山一角。我们现在还不知道,Drosha是怎么从细胞核跑到细胞质里的——它有核定位信号,是什么信号让它「叛逃」到细胞质?它的胞质异构体是否在不同组织里有不一样的功能?比如在肿瘤细胞里,Drosha的胞质分布和肿瘤的恶性程度相关,这是否和它调控脂肪生成的功能有关?
还有,Drosha在细胞质里除了和Sec31A结合,还会不会和其他蛋白互动,参与更多代谢通路的调控?它的RNA剪刀功能和脂肪管家功能之间,有没有什么协调机制?会不会在某些情况下,这两个功能会互相干扰?
这些问题都还没有答案,但有一点可以确定:Drosha不再是那个只懂剪RNA的「工具人」了。它就像一个身兼数职的多面手,在细胞里扮演着远比我们想象中更复杂的角色。
当我们把目光从基因的「经典功能」里跳出来,会发现细胞里的每一个分子,都可能藏着跨界的惊喜。Drosha从RNA剪刀到脂肪管家的转变,不仅让我们对脂肪代谢的调控有了全新的理解,更提醒我们:生命的复杂性,永远超出人类的预设。
分子无界,功能不止预设。 未来,当我们再面对代谢疾病的难题时,或许不会只盯着那些已知的脂肪调控因子,而是会想起那个在细胞质里和Sec31A握手的Drosha——那个被我们误解了十几年的「跨界选手」,可能正是打开新疗法大门的钥匙。