对抗知识焦虑,从看懂这条开始
App 下载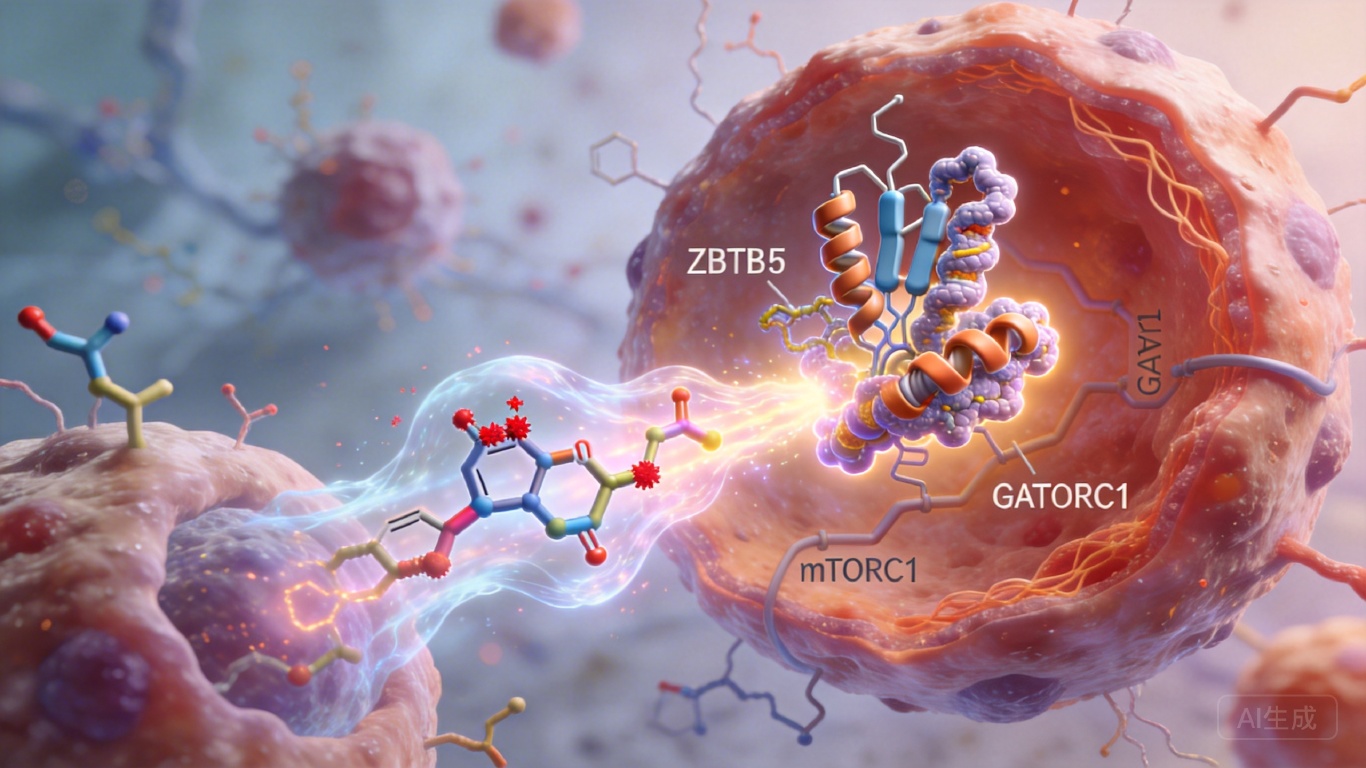
肿瘤靠“抢营养”躲免疫,新靶点打破耐药
肿瘤微环境|mTORC1通路|哈佛医学院|陆军军医大学|金凤实验室|肿瘤学|医学健康
对抗知识焦虑,从看懂这条开始
App 下载肿瘤微环境|mTORC1通路|哈佛医学院|陆军军医大学|金凤实验室|肿瘤学|医学健康
你或许听过免疫治疗能唤醒免疫系统杀癌细胞,但有近七成患者会出现耐药——不是免疫系统醒不过来,是它根本“饿到没力气干活”。肿瘤微环境里本就缺营养,癌细胞却还能在这种“饥荒”里疯狂抢食,甚至把免疫细胞的口粮也榨干。2026年,金凤实验室、陆军军医大学和哈佛医学院的联合团队,终于揪出了癌细胞这套“劫粮”操作的核心密码——一条被炎症信号劫持的营养感知通路。为什么癌细胞能在饿肚子时还能激活生长信号?这正是我们要解开的谜题。
要理解这套操作,得先搞懂细胞里的“营养总开关”——mTORC1。你可以把它想象成家里的电源总闸,只有当氨基酸这类“电费”充足时,闸才会合上,细胞才能启动生长代谢。正常情况下,一旦“电费”不足,GATOR1复合体就会自动跳闸,把mTORC1关掉,让细胞进入节能模式。
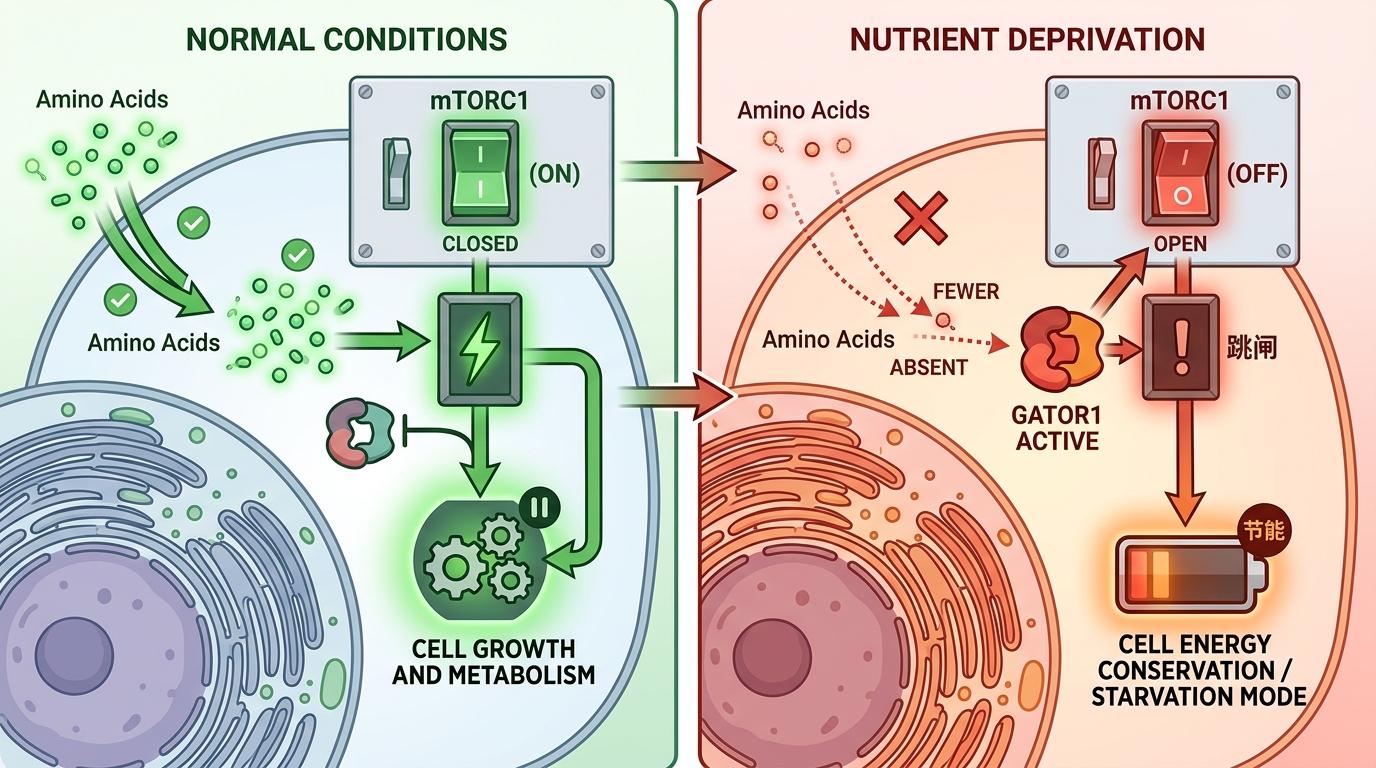
但癌细胞不遵守这个规则。
研究发现,肿瘤微环境里的炎症细胞因子,会给癌细胞里的ZBTB5蛋白“打个标记”——让它第127位丝氨酸发生磷酸化。这个磷酸化的ZBTB5就像个被收买的电工,它会找来Cullin3 E3泛素连接酶这个“拆迁队”,把负责跳闸的GATOR1复合体里的NPRL2给拆了。没了NPRL2,GATOR1就成了摆设,哪怕营养再匮乏,mTORC1这个总闸也会一直合上。
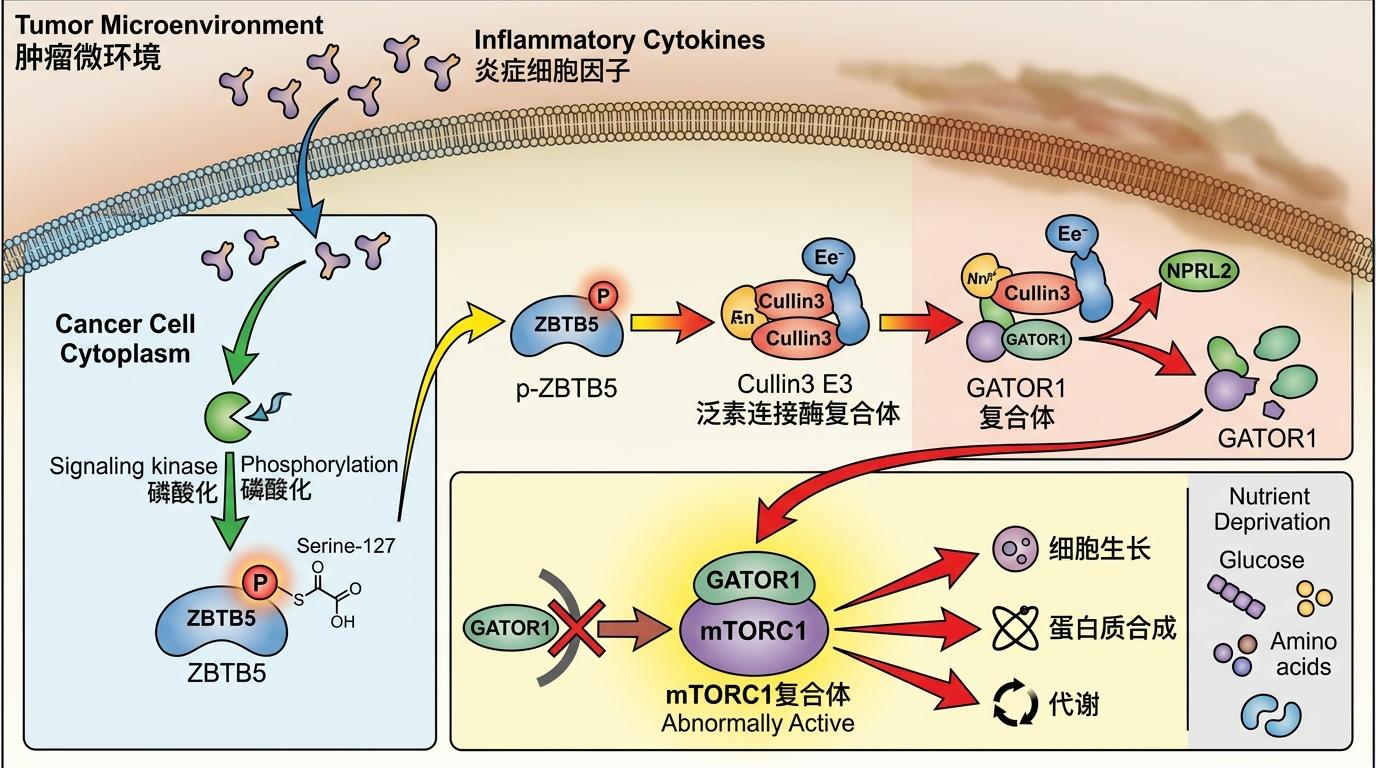
简单说就是:炎症信号→ZBTB5磷酸化→拆毁GATOR1→mTORC1持续激活→癌细胞疯狂抢氨基酸。
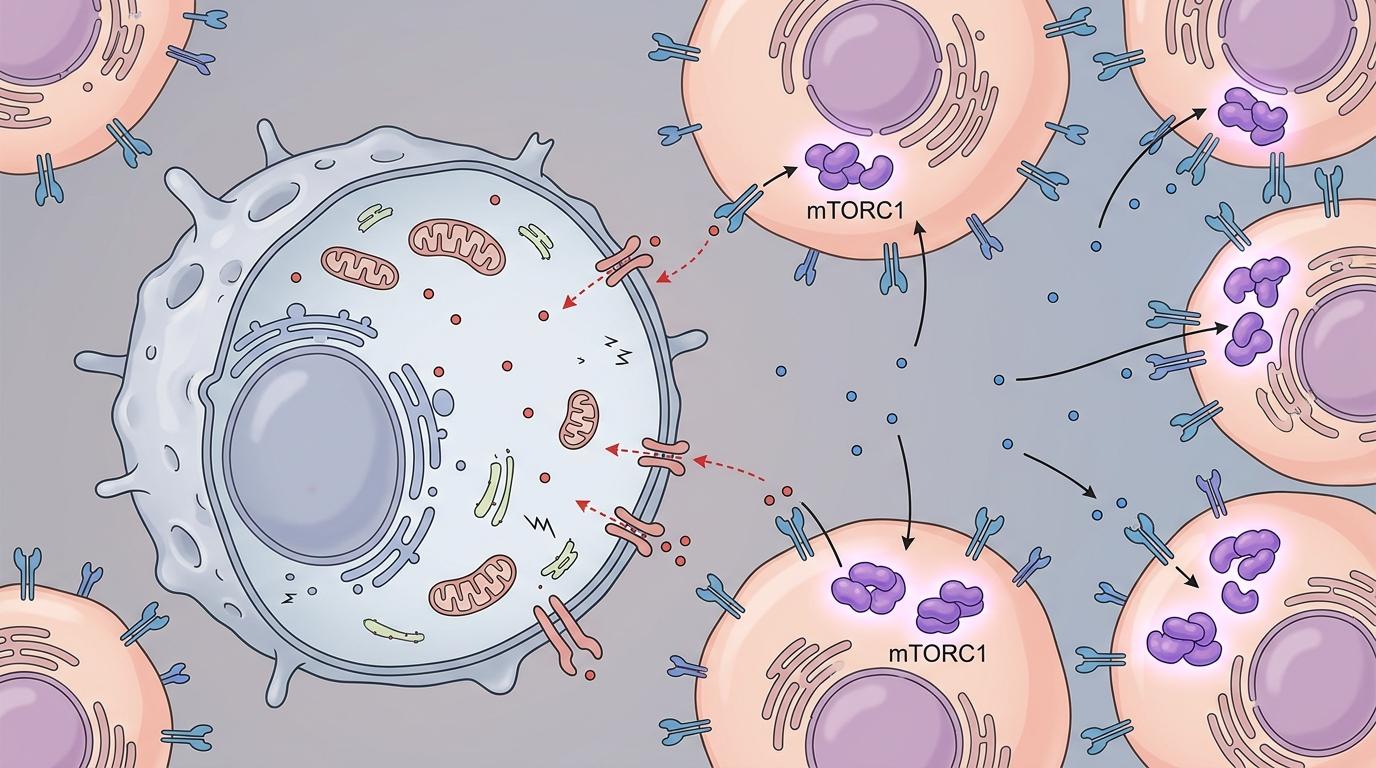
癌细胞抢来的氨基酸,不仅用来自己生长,还直接断了免疫细胞的活路。
CD8+T细胞是免疫系统杀癌细胞的主力军,但它同样需要氨基酸来维持活性。当癌细胞通过激活的mTORC1通路,把肿瘤微环境里的氨基酸几乎榨干时,CD8+T细胞就会陷入“营养饥荒”:要么代谢功能崩溃,要么直接饿死。这就解释了为什么免疫治疗会失效——不是T细胞不想干活,是它连“吃饭”的力气都没有了。
团队在细胞实验和小鼠模型里验证了这个逻辑:当他们用手段阻断ZBTB5的第127位丝氨酸磷酸化时,GATOR1复合体保住了,mTORC1的疯狂状态被按下暂停键,癌细胞的氨基酸摄取量骤降,肿瘤微环境里的氨基酸水平慢慢恢复。原本饿到濒死的CD8+T细胞活了过来,重新开始攻击癌细胞,就连对免疫检查点抑制剂耐药的肿瘤,也开始对治疗有了响应。
更关键的是,这套机制不仅解释了原发性耐药,还覆盖了获得性耐药——哪怕癌细胞后期进化出了耐药性,只要切断这条营养劫持通路,就能重新唤醒免疫反应。
当然,这个发现离真正的临床应用还有一段路要走。
首先是药物开发的难点:ZBTB5的第127位丝氨酸磷酸化位点,是个蛋白上的微小标记,要开发出能精准阻断这个位点的小分子药物,同时不影响正常细胞的mTORC1功能,难度不小——毕竟正常细胞也需要这个营养开关来维持代谢。
其次是肿瘤的异质性。不同肿瘤里的炎症水平、ZBTB5表达量都不一样,比如胰腺癌这类炎症水平极高的肿瘤,这条通路可能异常活跃,但一些炎症水平低的肿瘤,可能这套机制的作用就没那么强。未来还需要更多研究,来明确哪些患者最适合靶向这条通路的治疗。
不过值得期待的是,团队已经在小鼠模型里看到了明确的效果:阻断ZBTB5–pS127后,联合免疫检查点抑制剂,原本耐药的肿瘤缩小了近60%,CD8+T细胞的数量和活性都恢复到了接近正常水平。这至少给了我们一个清晰的方向——与其只盯着癌细胞杀,不如先把它抢营养的“脏手”按住。
过去我们总把肿瘤耐药归结为癌细胞的“狡猾”,但这次的研究让我们看到,所谓的“狡猾”,本质上是癌细胞利用了细胞本身的生存逻辑——它把炎症信号变成了“劫粮令”,把营养感知通路变成了“抢粮通道”。
饿不死的癌细胞,喂不饱的免疫细胞。 这句话精准戳中了肿瘤免疫逃逸的核心。而ZBTB5–pS127–mTORC1轴的发现,给我们提供了一个全新的解题思路:与其和癌细胞抢着杀癌细胞,不如先打破它对营养的垄断,让免疫系统重新获得“作战口粮”。
未来的癌症治疗,或许不再是单一的“杀敌”,而是一场关于“营养分配”的精准调控——我们要做的,就是把癌细胞多拿的那一份,还给本该拿到它的免疫细胞。