对抗知识焦虑,从看懂这条开始
App 下载
小脑学习的隐秘开关:抑制信号竟能促进学习
哈佛医学院Regehr团队|抑制性中间神经元|攀缘纤维|浦肯野细胞|小脑|神经生物学|生命科学
对抗知识焦虑,从看懂这条开始
App 下载哈佛医学院Regehr团队|抑制性中间神经元|攀缘纤维|浦肯野细胞|小脑|神经生物学|生命科学
当你学会骑自行车、打字或者记住一个新动作时,你的小脑正在进行一场精密的神经运算——这其中,攀缘纤维扮演着「教练」的角色:它通过强烈的电信号激活浦肯野细胞,触发树突内的钙信号,而这种钙信号正是小脑学习的核心密码。但这里藏着一个悖论:攀缘纤维同时还会激活一类抑制性中间神经元,按常理应该削弱浦肯野细胞的活性,阻碍学习。
哈佛医学院的Regehr团队花了数年时间,像侦探一样追踪每一个神经信号的走向,终于在2026年3月解开了这个矛盾——答案藏在两种看似功能相反的中间神经元里。
你可以把小脑的神经回路想象成一个精密的自行车刹车系统:浦肯野细胞是车轮,MLI1是直接捏紧刹车的手,而MLI2则是按住那只手的另一只手——也就是「去抑制」信号。
过去科学家以为所有中间神经元都是「刹车手」,直到Regehr团队用连续电子显微镜重建了神经连接:攀缘纤维确实会接触两种中间神经元,但和MLI2的接触点数量是MLI1的2.4倍,接触面积也大得多。就像给MLI2接了一根更粗的电线,攀缘纤维的谷氨酸信号会优先「溢出」激活MLI2。
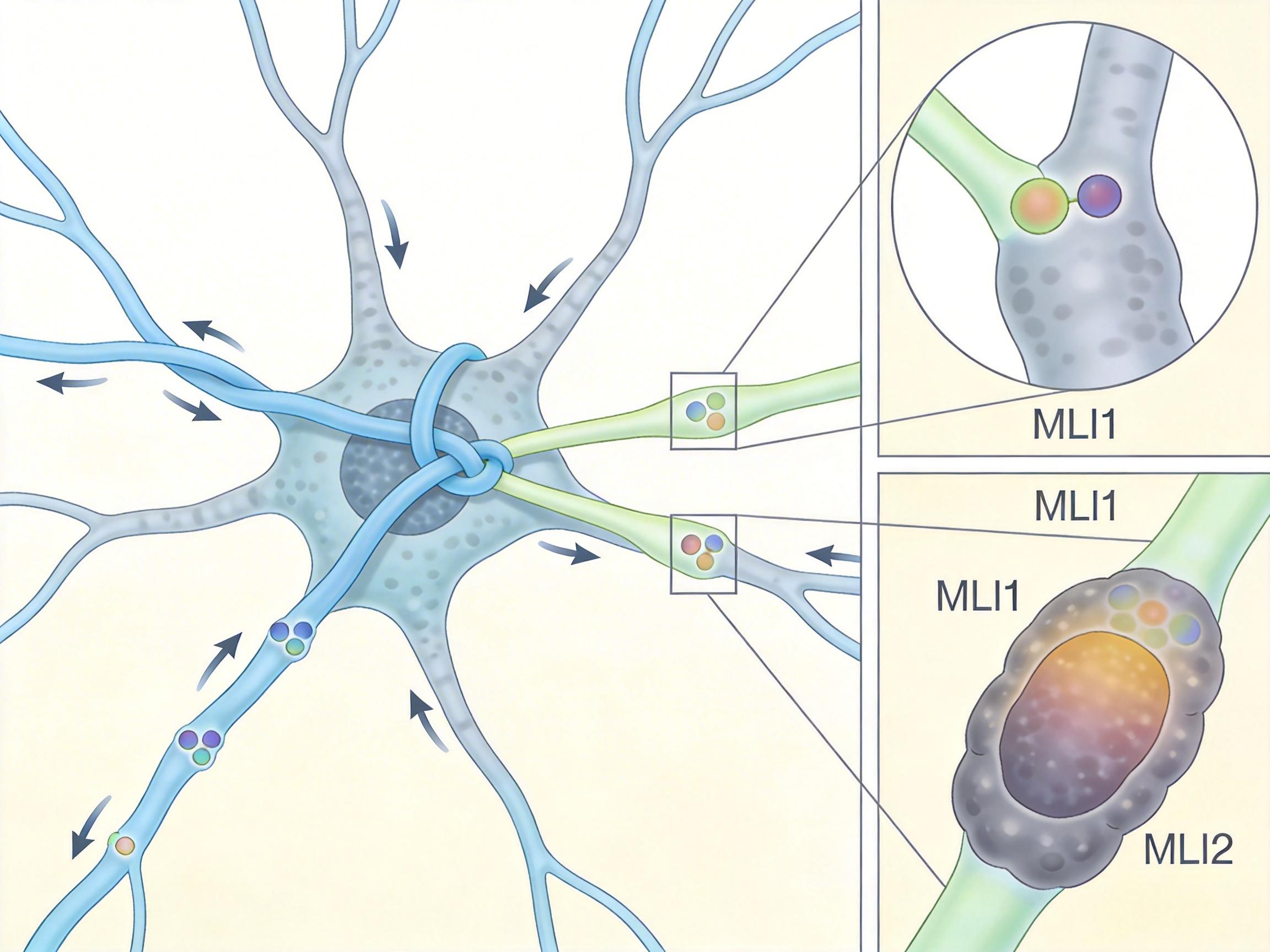
当攀缘纤维发出「学习指令」时,MLI2会先一步抑制MLI1,相当于按住了刹车手,让浦肯野细胞的钙信号不仅没被削弱,反而因为刹车被松开而变得更强。这就解释了为什么抑制性信号反而能促进学习——它抑制的不是学习本身,而是阻碍学习的力量。
单根攀缘纤维的信号已经能触发学习,但Regehr团队发现,当多根攀缘纤维同步激活时,浦肯野细胞的钙信号会大幅增强,学习效率也会显著提高。
他们在自由活动的小鼠脑中植入了神经像素电极,记录到了清晰的信号链条:自发的攀缘纤维活动只会轻微激活MLI2,抑制MLI1的效果有限;但当有外界感官刺激(比如吹向小鼠眼睛的气流)时,多根攀缘纤维会同步放电,MLI2的激活强度翻倍,MLI1则被彻底压制。此时浦肯野细胞的树突钙信号幅度增加了近30%,持续时间延长一倍——这正是触发长期突触抑制(LTD)、巩固学习记忆的关键。
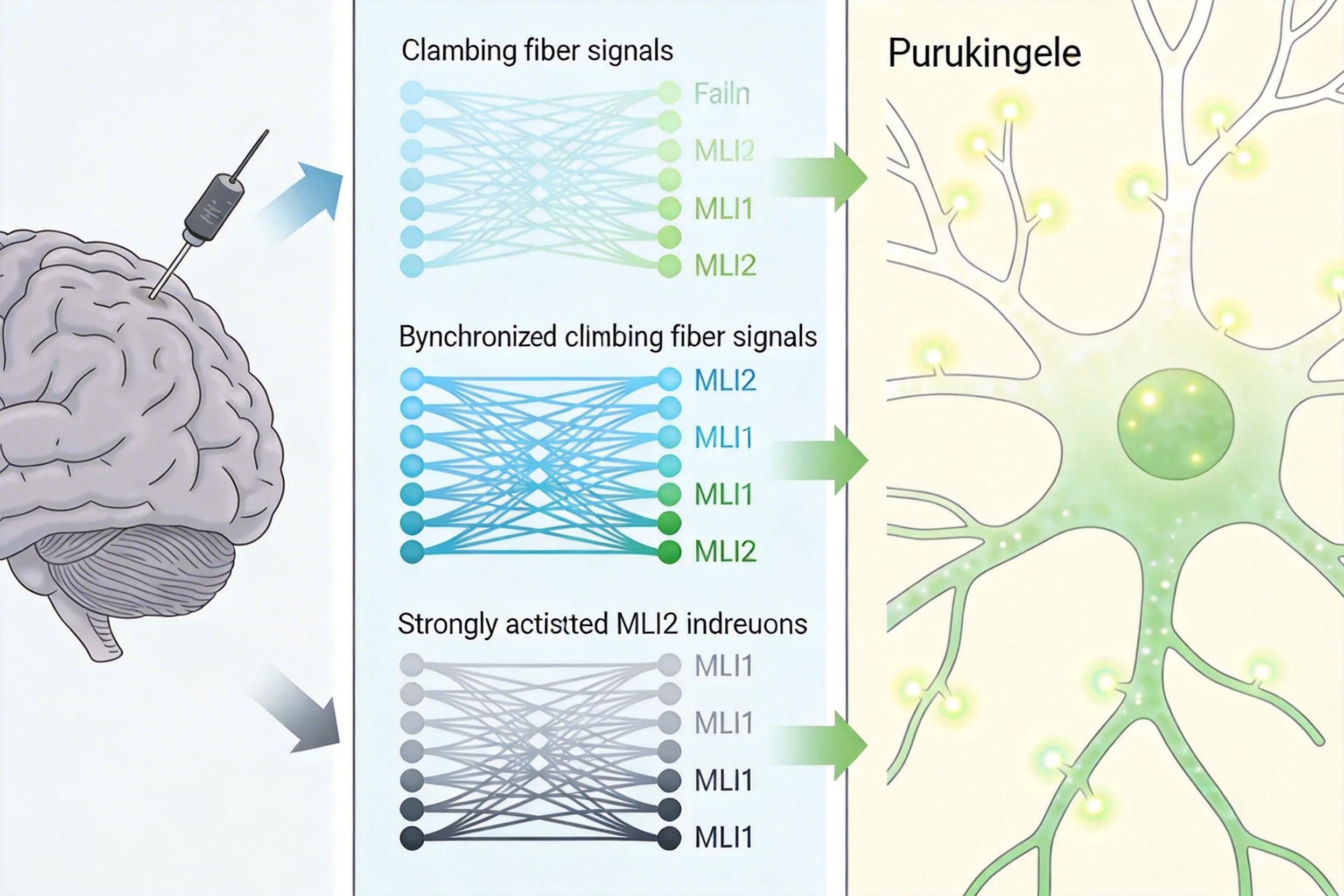
这种同步调控就像一群教练同时喊口令,能让浦肯野细胞更精准地捕捉到学习信号,避免被无关的抑制信号干扰。计算模型也验证了这一点:同步激活5根以上的攀缘纤维,就能让整个回路的学习效率提升2.7倍。
尽管回路机制已经清晰,但仍有一个关键问题悬而未决:攀缘纤维是如何在分子层面精准「选择」激活MLI2的?
目前的研究只发现MLI2表面有更多能结合谷氨酸的NMDA受体,且接触点附近的谷氨酸转运体更少,让谷氨酸更容易聚集。但具体是什么分子标记决定了这种选择性连接,还没有答案。Regehr团队推测,可能是神经粘附分子的差异——就像不同的锁对应不同的钥匙,MLI2表面有特殊的「钥匙孔」,能优先接收攀缘纤维的信号。
这个问题的答案不仅能深化我们对小脑学习的理解,还可能为治疗小脑共济失调、运动学习障碍提供新靶点:如果能人工激活MLI2,就能增强浦肯野细胞的活性,改善患者的运动协调能力。
从最初发现攀缘纤维的「教练」角色,到解开抑制信号促进学习的悖论,人类花了近半个世纪才看清小脑学习的核心回路。这背后是神经科学技术的迭代:从电子显微镜的精细重建,到体内多电极记录的实时追踪,再到计算模型的模拟验证。
「抑制不是阻碍,而是精准的调控」——这不仅是小脑的学习逻辑,也可能是大脑所有高级功能的共同密码。当我们下次学会一个新动作时,不妨想想:你的脑中正在发生一场由两种中间神经元主导的「刹车与松闸」游戏,而正是这种精密的平衡,让学习成为可能。