
10 天前
10 天前
你或许听过CRISPR——那个能精准编辑基因的“分子剪刀”,但很少有人知道,它最初是细菌用来对抗病毒的“微型免疫系统”。在这场持续了数十亿年的微观战争里,细菌靠一种叫环状寡腺苷酸(cOA)的“信号弹”启动防御。科学家已经破解了三种信号弹的密码,唯独对战场上产量最高的那种——cA5,始终找不到对应的“接收器”。就像发现了满场的信号,却不知道谁在指挥战斗。直到2026年3月,福州大学和西湖大学的联合团队,终于在显微镜下抓住了那个藏了亿万年的关键角色。
我们可以把细菌的免疫防线想象成一套密码锁:病毒入侵时,Cas10蛋白会根据入侵者的特征,打出不同组合的cOA“密码牌”——cA3、cA4、cA5、cA6,每一种密码对应一个专属的“锁孔”,也就是效应蛋白。过去十年,科学家找到了cA3、cA4、cA6的锁孔,唯独产量最高的cA5,像一把找不到锁的钥匙,在细胞里飘来飘去。
这次团队从放线菌里揪出的Csm6-2,就是cA5专属的锁孔。生化实验显示,它对cA5的响应效率是cA6的几十倍——哪怕两种信号弹和它的结合力度差不多。这就像两把钥匙都能插进锁孔,但只有对的那把能转动锁芯。
冷冻电镜给出了答案:当cA5钻进Csm6-2的CARF结构域,这个“锁孔”会像手掌一样完全合拢,紧紧裹住信号弹,同时触发另一端的HEPN结构域——也就是“剪刀”部分,开始疯狂切割病毒RNA;而cA6因为多了一个腺苷酸,像钥匙上多了个凸起,锁孔根本合不上,剪刀也就纹丝不动。
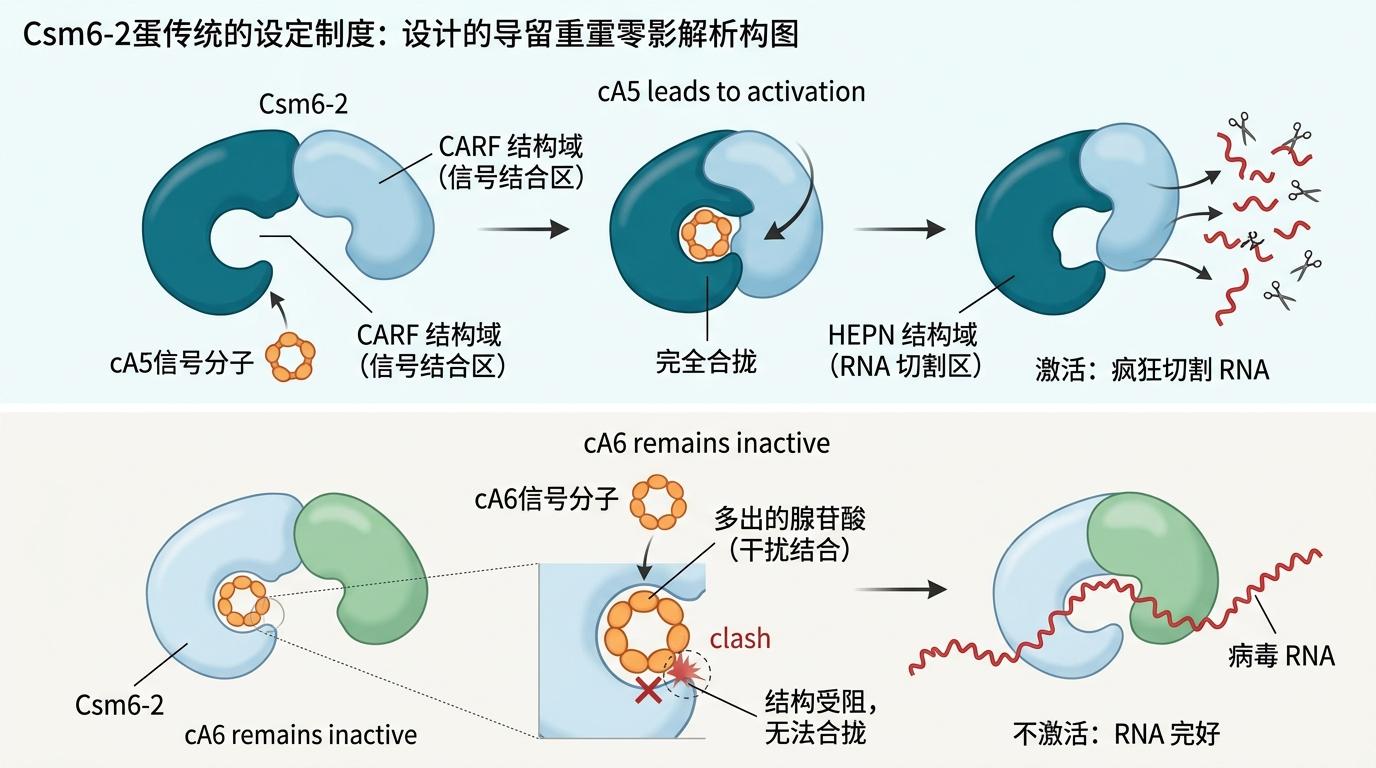
细菌的这套防御系统狠起来连自己都杀——非特异性切割RNA的剪刀一旦启动,连宿主自身的RNA也会被切碎,最后只能同归于尽。所以正常的Csm6蛋白都会在CARF结构域里自带“刹车”:激活一段时间后,就把手里的cOA信号弹降解掉,让剪刀停下来。
但Csm6-2偏不按常理出牌。它的CARF结构域是个“护短”的锁孔,一旦抓住cA5就死死护住,根本不会降解;真正的刹车藏在HEPN结构域里——这部分既是切割病毒RNA的剪刀,也是慢慢降解cA5的“消音器”。
这种设计的好处显而易见:它不会一下子把信号弹全清掉,而是留着一部分维持基础的“警戒状态”,既能及时响应新的病毒入侵,又不会因为过度反应把自己搞死。就像一支军队,打完仗不会立刻解散,而是保留常备军随时待命。
现在我们终于补全了III型CRISPR信号网络里缺失的一环,但这仅仅是开始。
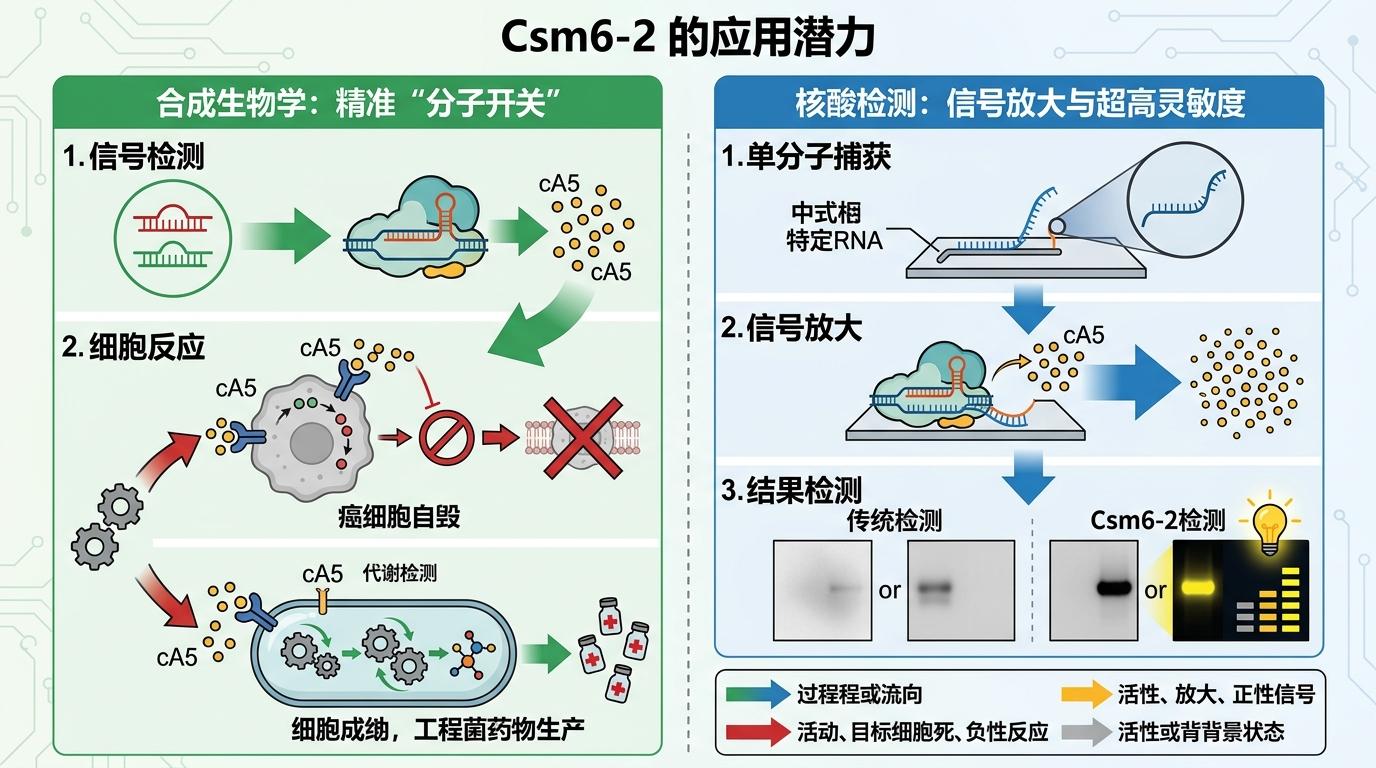
从应用层面看,Csm6-2对cA5的超高特异性,给合成生物学提供了一个精准的“分子开关”——我们可以用它来设计只对特定信号响应的细胞电路,比如让癌细胞在检测到特定RNA时启动自毁程序,或者让工程菌在特定环境下开始生产药物。在核酸检测领域,它的信号放大能力也可能让检测灵敏度再上一个台阶,甚至实现单分子级别的检测。
但更值得关注的是,这背后藏着细菌免疫的无限可能。我们现在只找到了几种cOA的效应蛋白,谁知道还有多少信号弹和锁孔藏在微生物的基因组里?那些能在极端环境下生存的古菌、深海里的细菌,说不定握着更精妙的防御机制——而这些,都是未来对抗超级病毒、开发新型抗菌策略的宝库。
当我们在实验室里破解Csm6-2的结构时,其实是在重读一本写了数十亿年的战争日记。每一个蛋白的折叠方式、每一次构象的变化,都是细菌和病毒在亿万年的厮杀里打磨出来的生存智慧。
微观战场的每一步,都是进化的奇迹。
我们现在能做的,就是把这些散落的智慧碎片捡起来,拼成更完整的图景。说不定下一个藏在微生物里的秘密,就能帮我们打赢一场人类自己的战争。
点击充电,成为大圆镜下一个视频选题!