对抗知识焦虑,从看懂这条开始
App 下载
染色质不是转录障碍,是扭矩缓冲器
康奈尔大学|角光学陷阱|扭矩缓冲|染色质结构|RNA聚合酶II|分子细胞生物学|生命科学
对抗知识焦虑,从看懂这条开始
App 下载康奈尔大学|角光学陷阱|扭矩缓冲|染色质结构|RNA聚合酶II|分子细胞生物学|生命科学
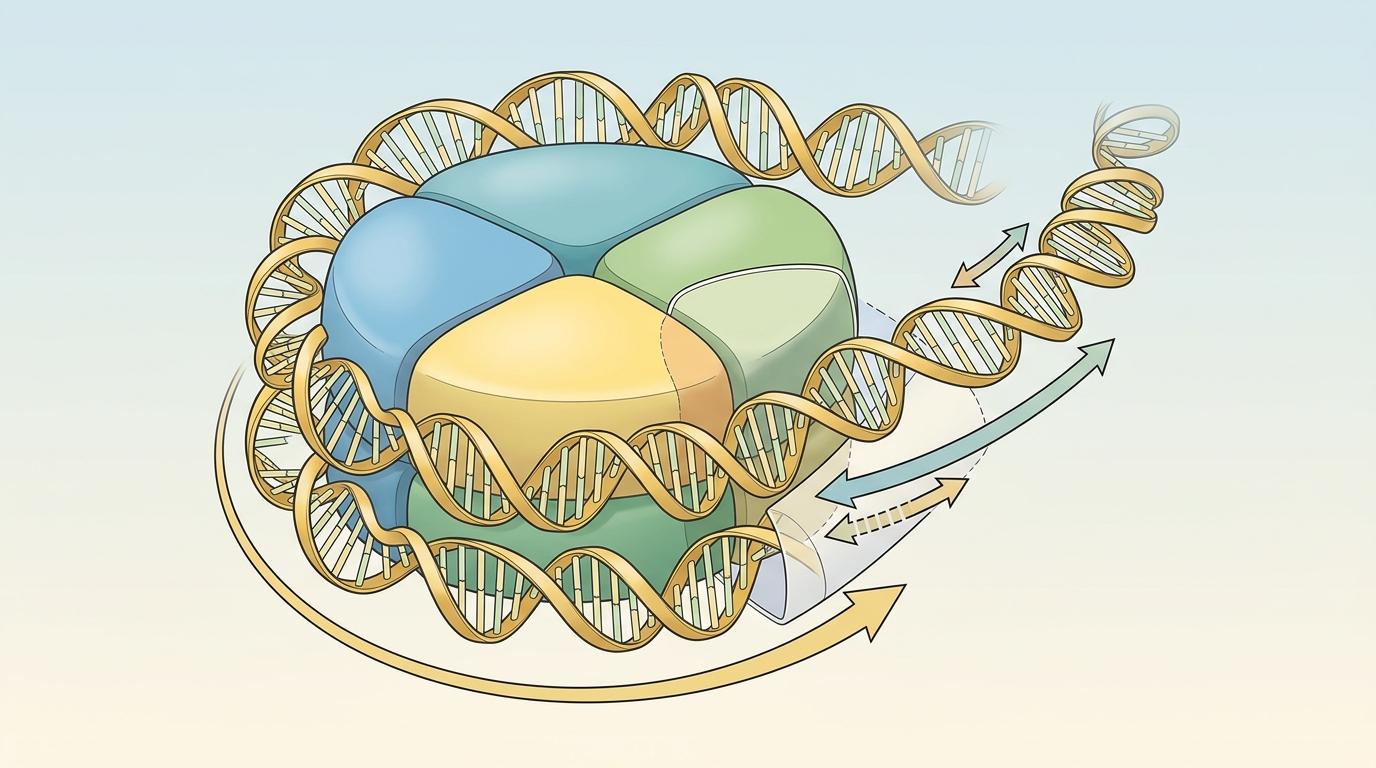
想象你是一个沿着螺旋楼梯向上跑的人——每跑10.5级台阶,你就得跟着楼梯转一整圈,还要在身前把台阶拧得更紧,身后的台阶却松得要散架。这就是RNA聚合酶II(Pol II)的日常:它沿着DNA的右手螺旋前进,必须同步旋转,在前方拧出正向超螺旋,像越来越紧的弹簧一样阻碍自己的路。过去我们一直以为,缠在组蛋白上的染色质只是它要推开的又一道墙。但康奈尔大学的科学家用镊子和激光盯住单个分子后发现,这道墙其实是个减震器。
你可以把Pol II理解成一个自带动力的钻头:它一边沿着DNA“钻”,一边得跟着DNA的螺旋转,这就会产生扭转力——也就是扭矩。用角光学陷阱盯着单个Pol II分子看,科学家测出它能拧出9皮牛顿·纳米的扭矩,差不多快能把DNA的双链拧开;如果加上转录因子TFIIS,这个数字会涨到13,相当于一个能撬动分子世界的旋转马达。
但问题来了,当Pol II在DNA上拧出越来越紧的正向超螺旋,它自己就会被这股力量卡住,甚至往回倒。这时候,过去被视为“路障”的染色质,反而显出了门道。
核小体是染色质的基本单位:DNA以左手螺旋的方式,绕在组蛋白上缠了1.8圈。这个左手螺旋刚好和DNA本身的右手螺旋“拧劲”相反——就像你在一根顺时针拧的绳子上,反方向绕了个圈,刚好能抵消一部分张力。
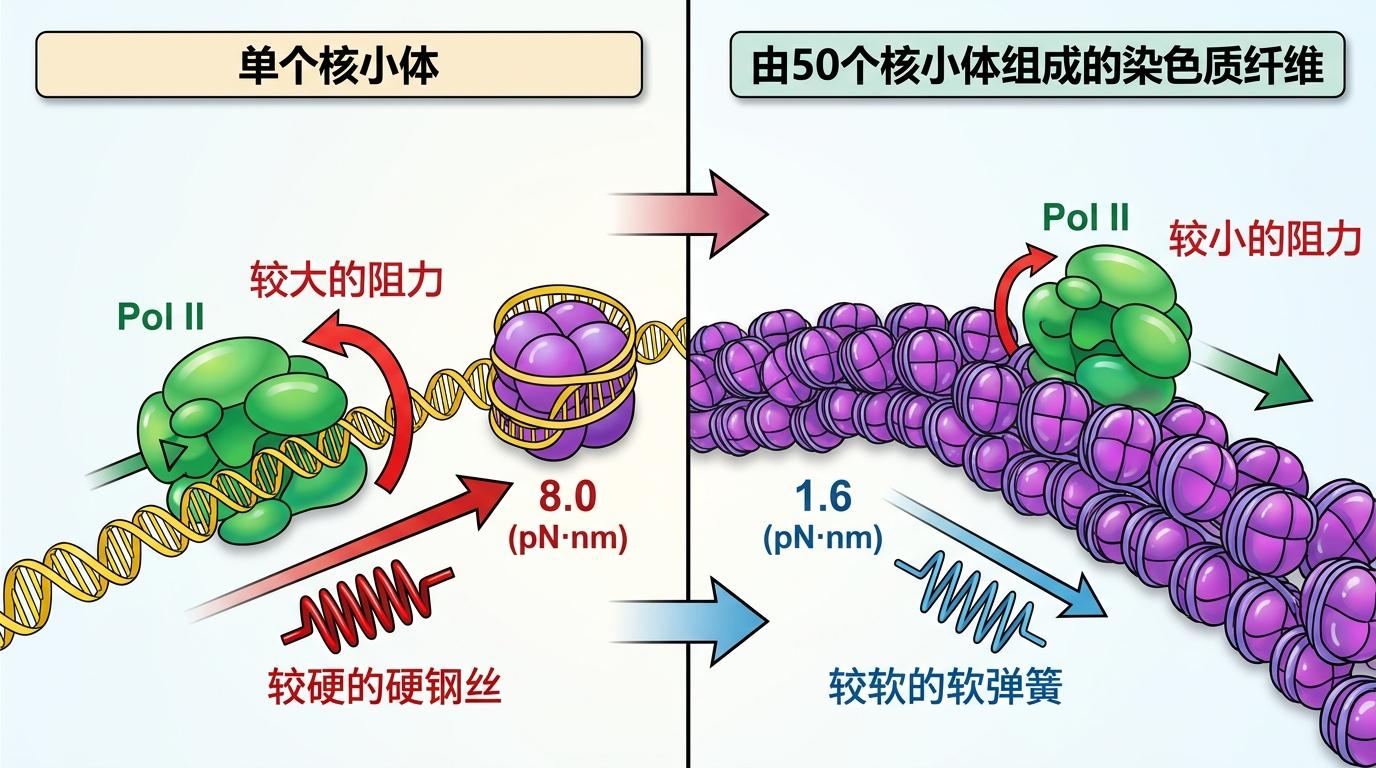
科学家用磁镊子做了个对比实验:让Pol II分别穿过单个核小体,和由50个核小体组成的染色质纤维。结果出乎意料:在完整染色质上,Pol II的通过率比单个核小体高得多。测算显示,裸DNA上的核小体给Pol II的阻力是8.0 pN·nm,而在染色质里,这个阻力降到了1.6 pN·nm——因为染色质的扭转刚度只有裸DNA的1/8,就像把硬钢丝换成了软弹簧,能把Pol II拧出来的扭矩“卸”掉大半。
当然,染色质的缓冲能力也有上限。当扭矩积累到超过它的承受范围,拓扑异构酶就会登场:它们像分子剪刀,把DNA链暂时剪开,释放掉多余的扭转应力,让Pol II能继续往前跑。实验里加了拓扑异构酶II后,Pol II一口气穿过了好几个核小体。
这些发现的背后,是单分子力学技术的突破。过去我们研究转录,只能看一群分子的平均表现,就像在电影院里看所有人的背影,不知道每个人在做什么。而角光学陷阱和磁镊子能抓住单个分子,给它加力、拧转,实时看它的反应——相当于把每个观众单独拉出来对话。
比如,科学家能精确控制Pol II遇到的扭矩大小,看它什么时候会卡住,什么时候能借助染色质的缓冲继续前进;也能观察TFIIS怎么帮Pol II“稳住方向”,不被扭矩逼得往回倒。这些过去只能靠猜的细节,现在都变成了能测量的数字。
不过这只是开始:我们还不知道染色质的更高阶结构,比如染色质环,怎么影响扭矩的传递;也不清楚组蛋白的化学修饰,会不会改变染色质的缓冲能力。这些问题,还得靠更精密的单分子技术来解答。
我们曾经把染色质当成基因表达的“绊脚石”,觉得它只是用来把DNA打包塞进细胞核的容器。但这次的发现像一把钥匙,打开了一个新的视角:生命的运转不只是化学信号的传递,还有力学力量的平衡——Pol II的扭矩、染色质的缓冲、拓扑异构酶的调节,共同构成了一个精密的力学系统。
“生命的秩序,藏在力的平衡里。” 当我们开始用力学的眼光看细胞里的分子,那些过去被忽略的细节,正在拼成一幅更完整的生命图景。而这不仅能帮我们理解基因怎么工作,更可能为癌症等疾病的治疗,找到全新的靶点——毕竟,很多疾病的根源,就是这个力学系统出了错。