对抗知识焦虑,从看懂这条开始
App 下载
细菌丢了“螺旋桨”,照样能跑能拐弯
鞭毛基因敲除|分子齿轮箱|黄杆菌|沙门氏菌|大肠杆菌|微生物组学|生命科学
对抗知识焦虑,从看懂这条开始
App 下载鞭毛基因敲除|分子齿轮箱|黄杆菌|沙门氏菌|大肠杆菌|微生物组学|生命科学
你大概以为,细菌就像微型摩托艇,全靠尾巴上那根旋转的鞭毛——相当于螺旋桨——才能到处跑。要是把这根螺旋桨拆了,它们就只能在原地等死?
亚利桑那州立大学的实验室里,这个被默认了几十年的常识,被一群无鞭毛的大肠杆菌和沙门氏菌推翻了。在铺了含糖琼脂的培养皿里,这些本该瘫痪的细菌,居然慢悠悠地在表面扩散开来,像被细流推着的落叶。更离谱的是另一类黄杆菌,它们压根就没长过鞭毛,却靠一套分子齿轮箱,像雪地摩托似的在表面滑行,还能随时倒车。
它们到底是怎么做到的?
Navish Wadhwa团队最初做的是个“负对照”实验:把大肠杆菌和沙门氏菌的鞭毛基因敲除,想看看失去动力的细菌会怎么“躺平”。结果培养皿里的菌落,居然在48小时内向外扩展了整整一厘米。
研究人员盯着显微镜看了无数小时,终于揪出了背后的机制:这些细菌在靠代谢“划船”。当它们啃食琼脂里的葡萄糖、麦芽糖时,会发酵出乙酸、甲酸这些酸性产物。这些产物在菌落边缘堆积,形成了局部的高渗透压区——相当于在菌落周围拉了个无形的“抽水机”,把琼脂里的水分吸过来,在菌落前沿鼓出一层几十微米高的液膜。这层液膜会慢慢向外流动,就像地表的细流,推着边缘的细菌群体一起前进。
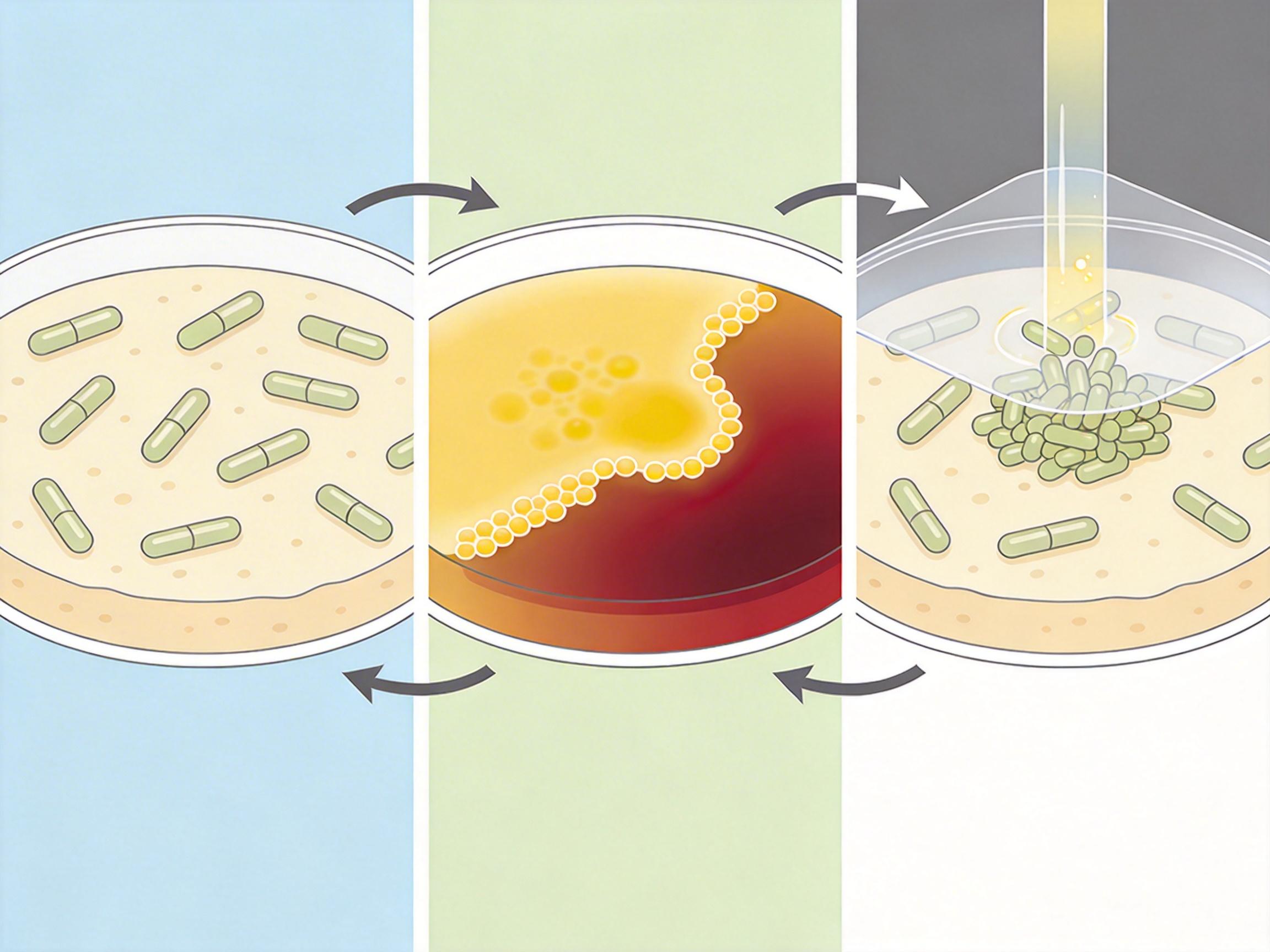
这种完全不依赖鞭毛的运动,被命名为“swashing”(摇荡运动)。它和细菌靠鞭毛集体冲锋的“群游”完全不是一回事:表面活性剂能抑制swashing,却会帮群游的细菌滑得更快;只要切断细菌的糖源,或者调高环境pH阻断发酵,swashing就会立刻停止。
最关键的是,这种“糖水动力”不需要细菌费力气“划桨”,全靠环境里的糖和自己的代谢本能。这也解释了为什么医院的导管、伤口的黏液里,明明没有鞭毛的致病菌也能偷偷扩散——这些地方,恰恰是含糖的湿润环境。
如果说swashing是细菌的“躺赢式”运动,那黄杆菌的滑行就是“硬核机械流”。这类细菌天生没有鞭毛,却靠一套叫T9SS(第九型分泌系统)的装置,在表面跑得飞快。
你可以把T9SS想象成一台贴在细菌体表的履带式雪地车:它的核心是一个由GldK蛋白组成的环形“齿轮”,内膜上的GldLM复合体像马达,靠质子动力推动齿轮旋转;而GldJ蛋白就是“换挡器”——当它的C端被截断时,齿轮会瞬间从逆时针旋转变成顺时针,细菌的运动方向也跟着反转,就像雪地车突然倒车。
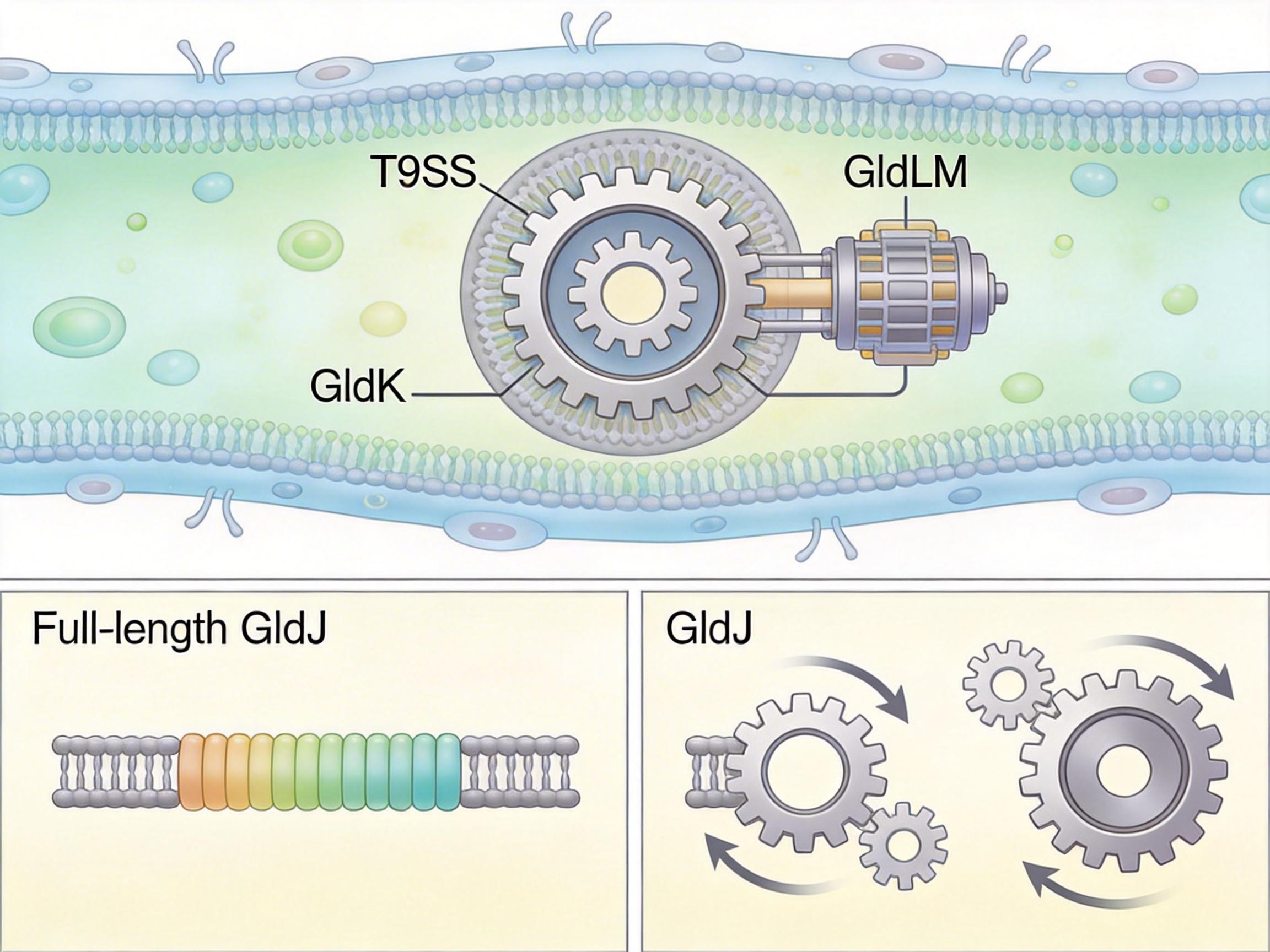
这套系统不止管运动,还兼职“快递员”:它会把黏附蛋白SprB送到履带上,让细菌能牢牢粘在表面,同时把代谢产物、毒力因子分泌到环境里。在口腔里,产黑普雷沃菌靠T9SS分泌的蛋白酶引发牙龈炎;而在肠道里,某些有益菌靠它分泌的蛋白保护抗体,帮你增强免疫力。
研究团队用冷冻电镜拍下了这套齿轮箱的细节:GldJ就像卡在齿轮缝隙里的拨片,稍微变个形状,就能改变整个机器的转向。这种精度,连人类的微型机械都很难做到。
这两个发现凑在一起,指向了一个被忽略的事实:细菌的“逃生工具箱”,比我们想象的要满得多。
过去我们对付细菌感染,总想着破坏它们的鞭毛——比如用药物让鞭毛转不动,或者干脆不让它们长鞭毛。但swashing和T9SS的存在证明,就算没了鞭毛,致病菌照样能在你的伤口里扩散,在导管上定植。更麻烦的是,这些“备用动力”往往和细菌的代谢、分泌系统绑定,比单一的鞭毛更难被针对性阻断。
但这也给了我们新的思路:既然swashing靠的是糖和发酵,那在医院的医疗器械表面,能不能降低糖含量、调高pH,断了它们的“燃料”?既然T9SS的齿轮箱离不开GldJ蛋白,能不能设计小分子药物,把这个“换挡器”卡死?
我认为,最关键的转变是:我们不能再只盯着细菌的“动力器官”,而要学会“断粮”和“拆机器”——从控制环境入手,从破坏它们的核心代谢和分子装置入手。毕竟,细菌能进化出备用动力,我们也能进化出更聪明的防控策略。
当我们以为已经摸清了细菌的所有花招时,它们总能掏出新的“底牌”。这两场发生在培养皿里的发现,其实是在提醒我们:微生物的世界,从来都不是按照人类的常识运转的。
“细菌的生存智慧,远超人类的想象。”这句话不是夸张——从靠糖水推自己走的大肠杆菌,到装着分子齿轮箱的黄杆菌,每一种看似“简陋”的生命,都藏着经过亿万年打磨的生存策略。
未来的抗菌战场,或许不再是我们和细菌的“武器竞赛”,而是一场关于“读懂对手”的博弈。毕竟,你得先知道敌人有多少种逃跑的办法,才能真正拦住它们。