对抗知识焦虑,从看懂这条开始
App 下载
95%细胞能量钥匙藏在线粒体,耶鲁找到开门密码
脂肪酸氧化|三羧酸循环|维生素B5|线粒体|辅酶A|分子细胞生物学|生命科学
对抗知识焦虑,从看懂这条开始
App 下载脂肪酸氧化|三羧酸循环|维生素B5|线粒体|辅酶A|分子细胞生物学|生命科学
你每天吃下的维生素B5,会在细胞里变成一把「能量钥匙」——辅酶A(CoA),它是细胞内100多种代谢反应的「通行证」,从分解食物到产生ATP都离不开它。更关键的是,人体里95%的辅酶A都集中在线粒体里——这个被称为细胞「能量工厂」的小结构,要靠它驱动三羧酸循环、脂肪酸氧化这些核心产能过程。但半个多世纪以来,科学家一直搞不懂一件事:辅酶A明明是在细胞质里合成的,它是怎么穿过线粒体那层不通透的双层膜,精准抵达「工厂车间」的?这就像你在公司楼下做好了关键零件,却找不到能送进核心机房的电梯。
要追踪辅酶A的行踪,首先得解决一个技术难题:它在细胞里几乎从不「裸奔」。作为一种辅因子,辅酶A总喜欢和其他分子绑定,形成结构各异的「辅酶A共轭物」——就像钥匙总插在不同的锁孔里,你很难找到一把单独的钥匙。过去的检测方法只能盯着某几种共轭物,永远拼不出完整的转运路线。
耶鲁大学的沈宏瑛(Hongying Shen)团队换了个思路:他们开发出一套结合稳定同位素标记的质谱技术,能同时捕捉细胞内33种辅酶A共轭物,还能精准区分出线粒体内的23种。就像给每个「带钥匙的锁」都贴了定位标签,终于能看清它们在细胞内的分布地图。
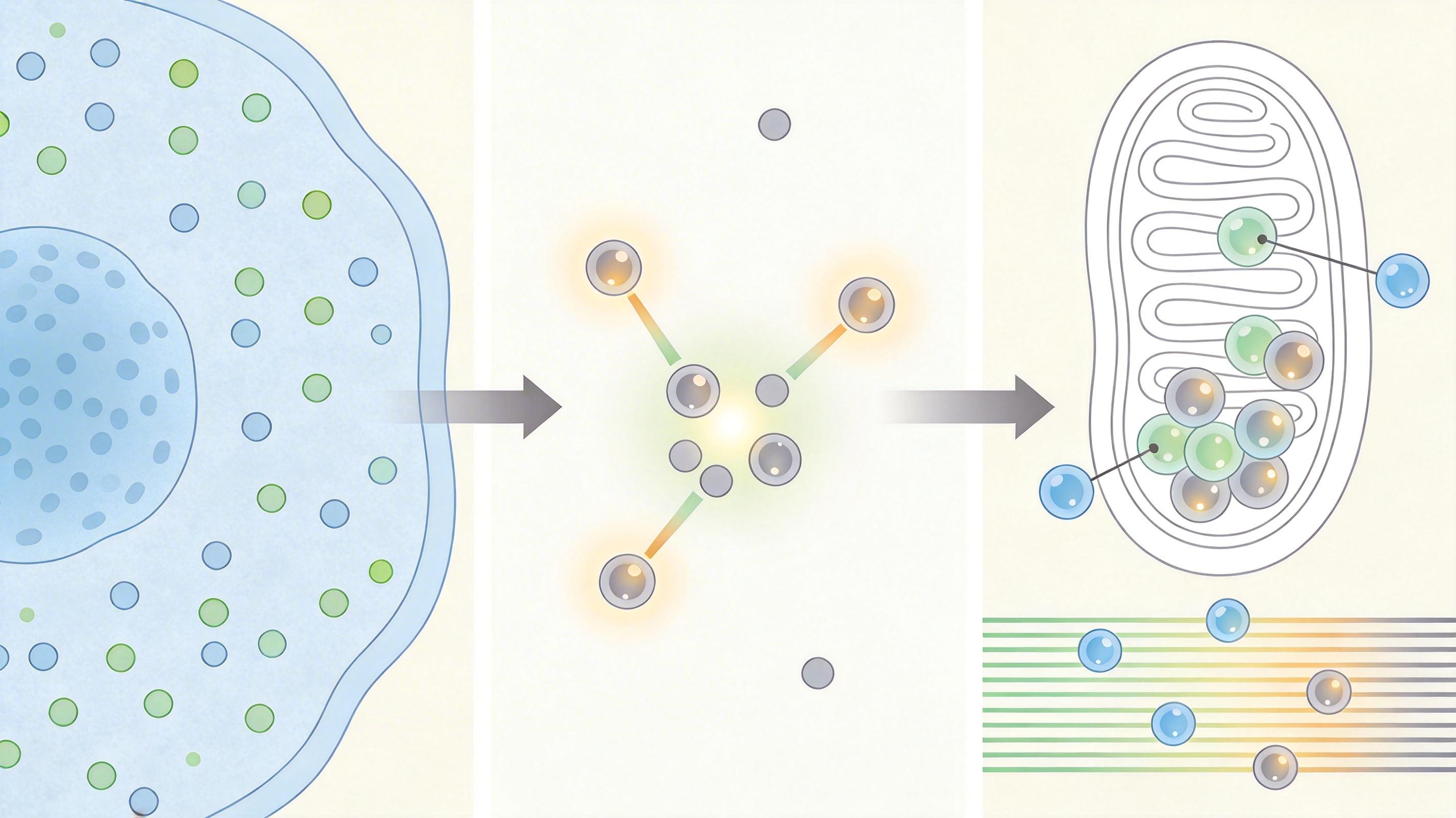
实验结果给出了第一个关键线索:线粒体内的辅酶A合成酶含量极低,只占细胞质的10%,根本不足以维持线粒体里95%的辅酶A浓度。
答案指向唯一的可能:辅酶A是从细胞质「运」进来的。
接下来的问题是,谁是负责运输的「电梯工」?团队用CRISPR基因编辑技术,逐一敲除了线粒体内膜上的候选转运蛋白,终于锁定了两个关键角色:SLC25A16和SLC25A42。
你可以把这两种蛋白想象成线粒体膜上的两个双向闸门——它们靠「反向交换」机制工作:把细胞质里的辅酶A送进线粒体的同时,把线粒体里的ADP或AMP运出去,既保证了分子的定向运输,又维持了膜两侧的电荷平衡。
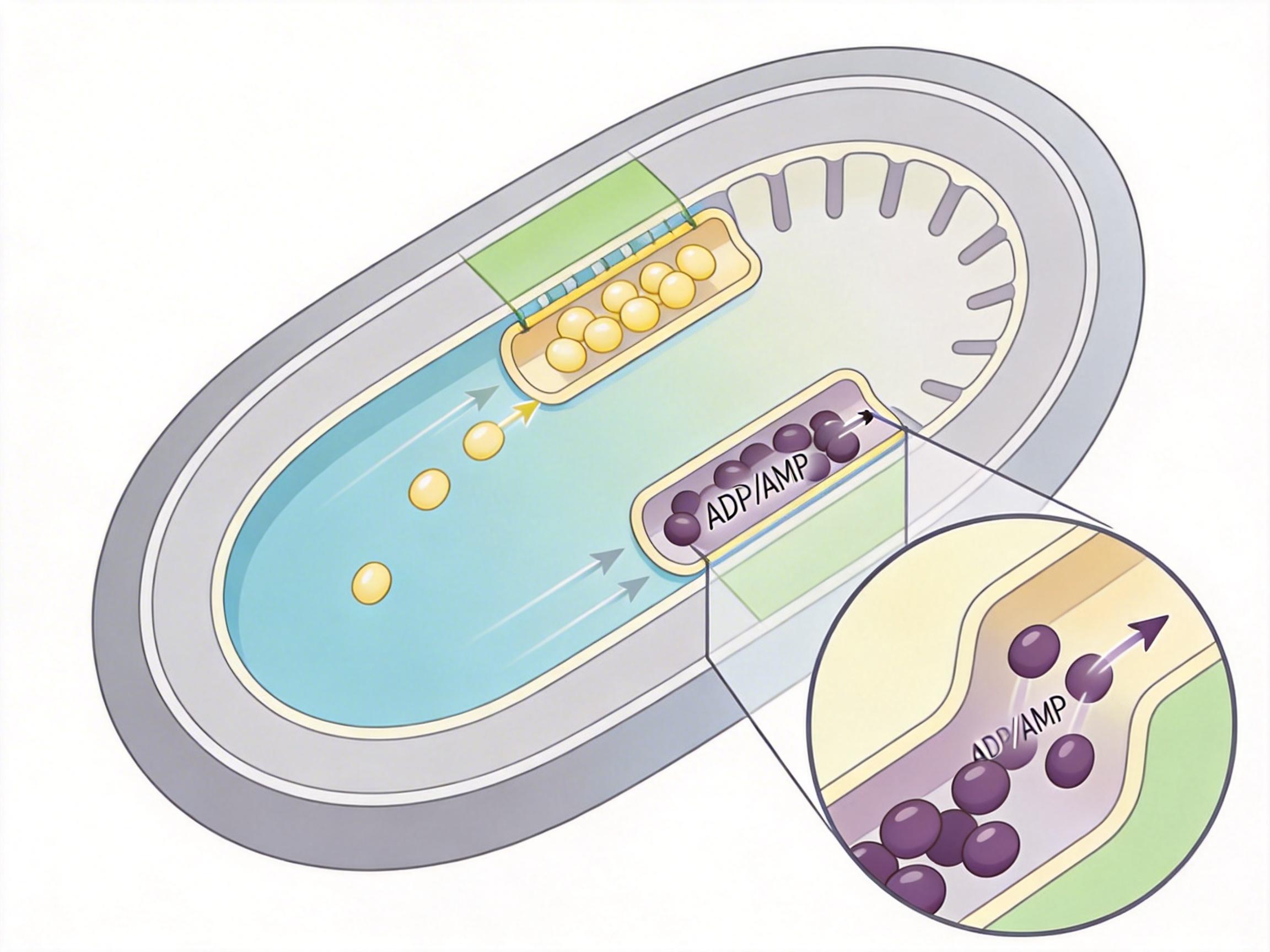
更有意思的是,这两个蛋白是「双保险」:单独敲除其中一个,线粒体内的辅酶A只下降一小部分,细胞还能勉强运转;但同时敲除两者,线粒体内的辅酶A含量直接暴跌90%以上——三羧酸循环停摆,脂肪酸氧化中断,细胞的能量供应近乎崩溃,增殖能力直接丧失。
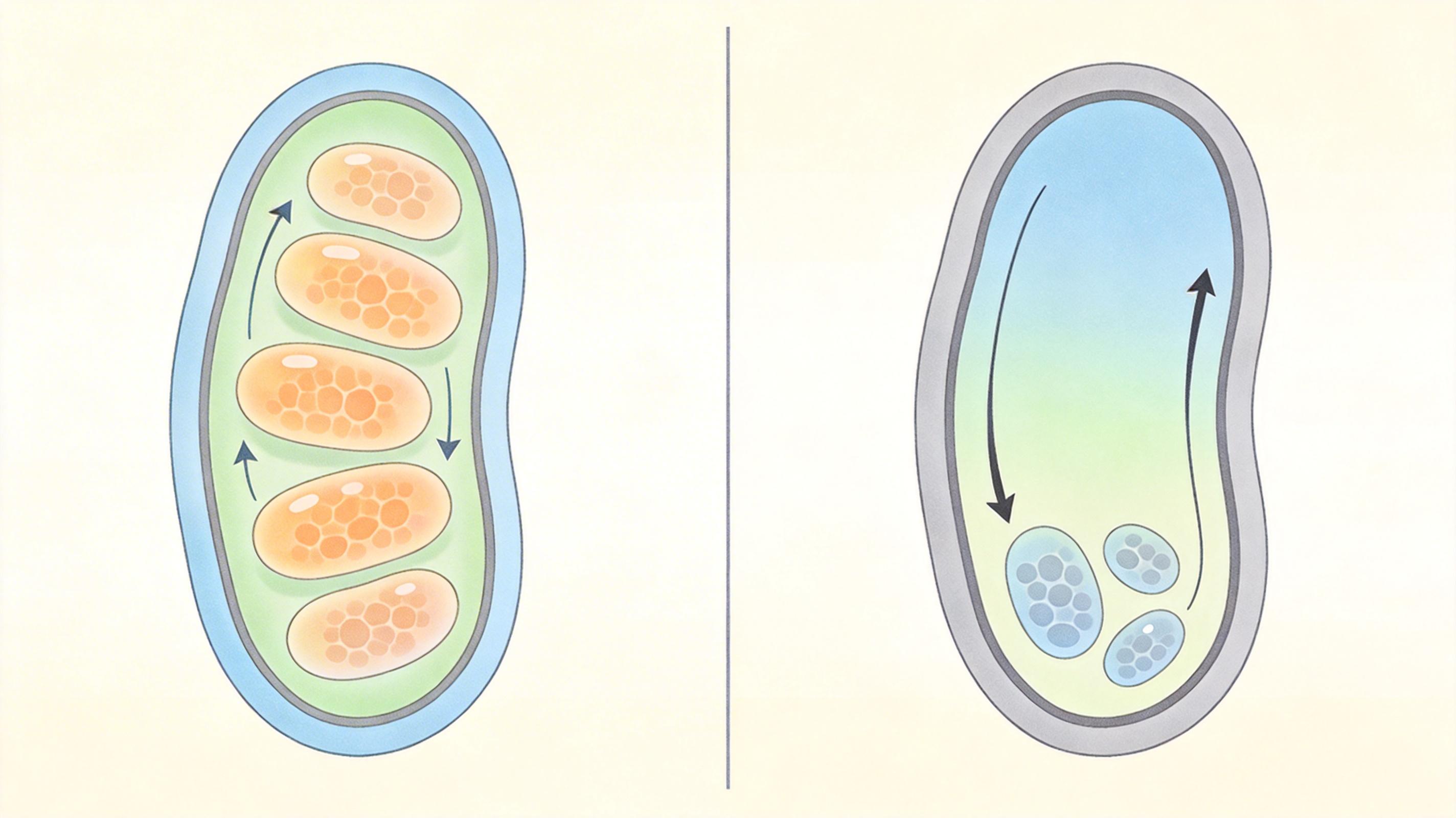
团队还在酵母实验中验证了这个机制:把人类的SLC25A42基因导入缺乏线粒体辅酶A转运蛋白的酵母突变体,酵母居然恢复了在非发酵碳源上的生长能力——这相当于给一辆没油的车重新接上了油管。
我认为,这个「双保险」设计是被媒体忽略的关键:它解释了为什么单一转运蛋白的突变只会引发特定疾病——比如SLC25A42的N291D突变会导致线粒体脑肌病,患者出现发育迟缓、癫痫等症状,但不会立刻致命。
搞清楚转运机制,不止是填补了教科书里的空白——它直接为一类罕见病打开了治疗的新方向。
比如PKAN(泛酸激酶相关神经退行性疾病),这种病是因为辅酶A合成的关键酶PANK2突变,导致脑内辅酶A不足,患者会出现运动障碍、脑内铁沉积等症状。过去的治疗思路是补充辅酶A前体,但这些分子很难穿过血脑屏障。现在我们知道,只要能激活脑内的SLC25A16和SLC25A42,就能让更多细胞质里的辅酶A进入神经元的线粒体,相当于给大脑的能量工厂开了更大的进料口。
沈宏瑛团队已经在研究神经元里的辅酶A转运调控机制——毕竟神经元是对能量需求最高的细胞之一,线粒体辅酶A的不足,可能是阿尔茨海默病、帕金森病等神经退行性疾病的共同诱因。
当然,目前的研究还有局限:我们还不知道这两个转运蛋白在不同组织里的调控差异,比如心脏和肝脏里的「闸门开关」是不是有不同的信号控制。但至少,我们终于找到了那台连接细胞质和线粒体的「电梯」,剩下的就是搞清楚怎么控制它的开关和速度。
从1910年耶鲁科学家发现维生素B复合体,到今天找到辅酶A进入线粒体的转运机制,这所大学在代谢研究上的传承已经延续了116年。这一次的发现,本质上是人类在细胞的微观世界里,又破解了一个维持生命运转的「基本程序」。
每一个代谢细节的补全,都是在为生命的修复铺路。未来当医生面对那些因能量代谢紊乱引发的疾病时,他们不会再只盯着「能量工厂」本身,而是会想起那两个藏在膜上的「守门人」——正是它们,决定了细胞能不能拿到足够的「能量钥匙」,驱动生命的齿轮继续转动。