对抗知识焦虑,从看懂这条开始
App 下载
酵母极小着丝粒起源揭晓:垃圾DNA变核心结构
马克斯·普朗克研究所|着丝粒悖论|垃圾DNA|点着丝粒|酿酒酵母|分子细胞生物学|生命科学
对抗知识焦虑,从看懂这条开始
App 下载马克斯·普朗克研究所|着丝粒悖论|垃圾DNA|点着丝粒|酿酒酵母|分子细胞生物学|生命科学
当你啃着面包、抿着啤酒时,绝不会想到嘴里的酿酒酵母藏着一个困扰科学家40年的谜题:它的着丝粒——细胞分裂时负责拉扯染色体的「分子锚点」——只有人类着丝粒的百万分之一大。这种极致精简的「点着丝粒」,和绝大多数生物那动辄数百万碱基对的「区域着丝粒」截然不同,功能却丝毫不差。更诡异的是,明明是维持生命的核心结构,着丝粒的DNA序列却进化得比基因组其他区域快得多,这就是让学界挠头的「着丝粒悖论」。直到2026年,德国马克斯·普朗克研究所和纽约大学的联合团队,终于找到了连接这两种极端结构的缺失环节——但答案却颠覆了我们对「垃圾DNA」的认知。
你可以把着丝粒想象成染色体上的「挂扣」,细胞分裂时,纺锤丝就勾着这个挂扣把染色体拉向两个子细胞。人类的挂扣是一大块缠满线团的重复DNA,而酿酒酵母的挂扣却只有120个碱基对——差不多就是把人类的挂扣压缩成了一颗纽扣。
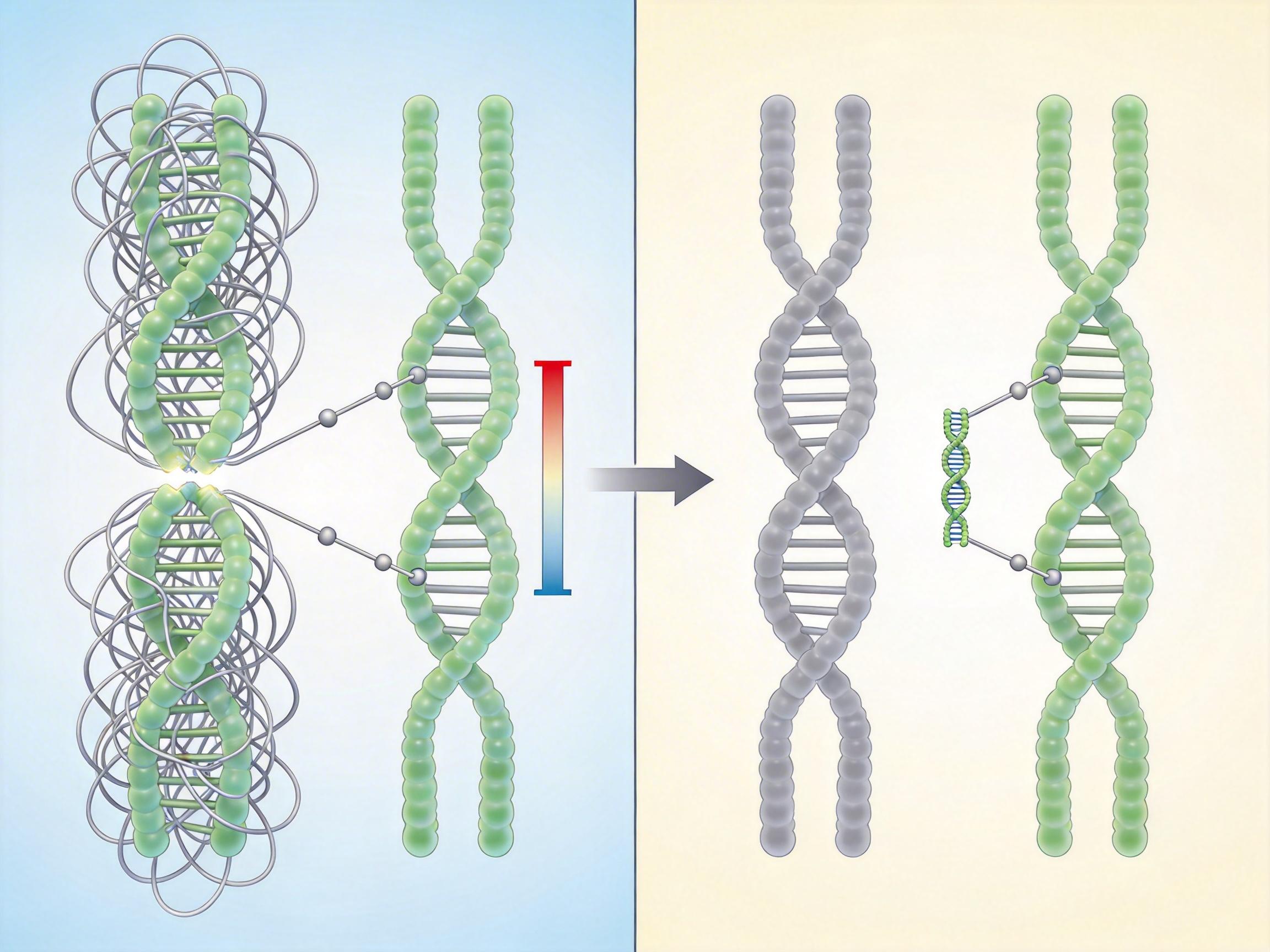
研究团队比对了200多种酵母的基因组,在酿酒酵母的近亲物种里找到了一种「半成品挂扣」:它们的着丝粒既不像人类那样臃肿,也不如酿酒酵母精简,核心序列里嵌着大量Ty5型逆转录转座子的片段——这就是曾被我们当成「基因组垃圾」的跳跃基因。
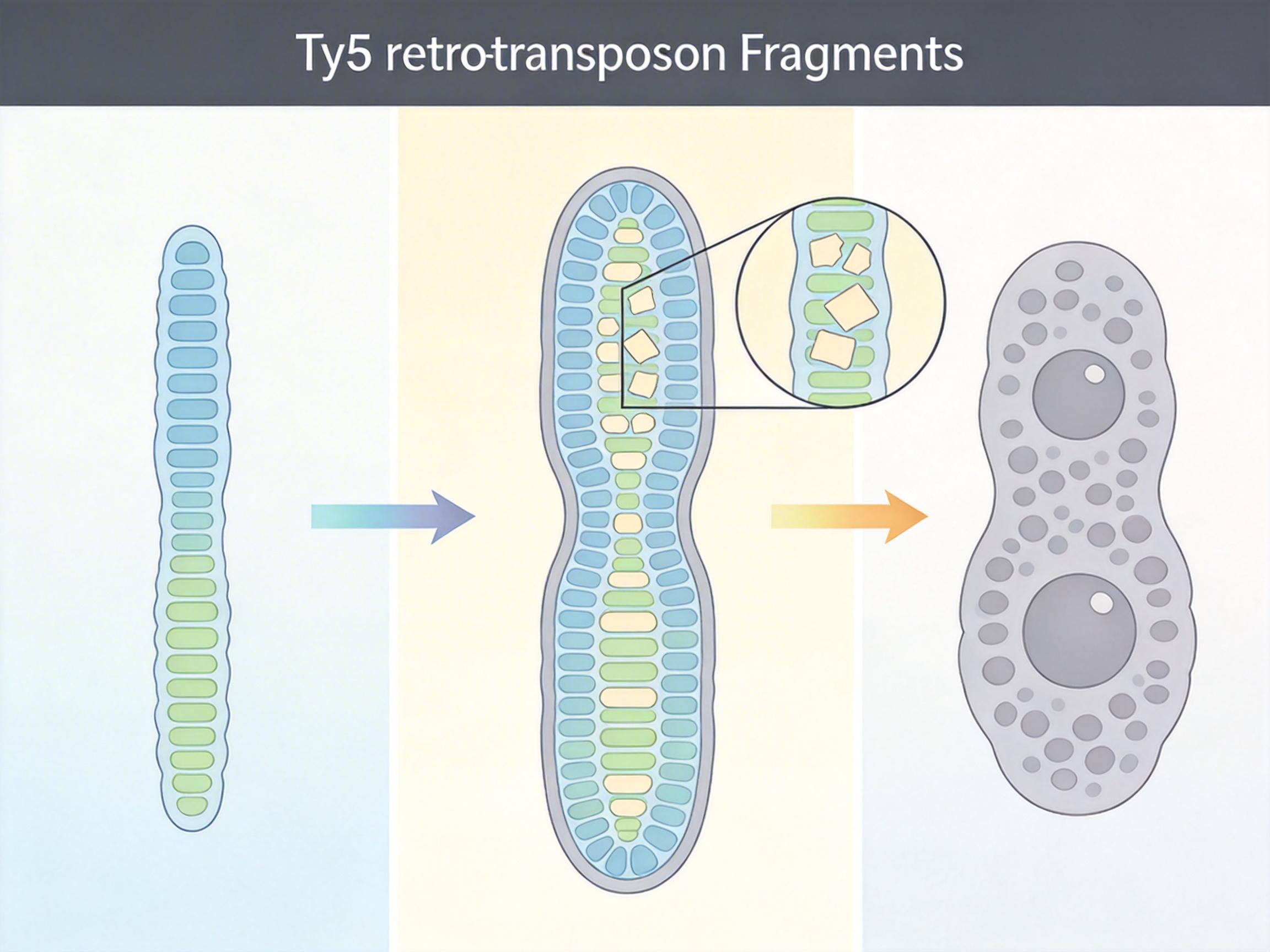
这些转座子原本是寄生在基因组里的「自私分子」,只会不断复制自己,看起来对细胞毫无用处。但在大约2.5亿年前,酿酒酵母的祖先丢失了传统的异染色质调控机制,原本用来稳定着丝粒的「分子胶水」没了。就在这时,那些Ty5转座子的序列刚好能被着丝粒蛋白复合体识别——自然选择抓住了这个偶然,把寄生的垃圾DNA,一步步改造成了精准的点着丝粒。
着丝粒的功能必须绝对稳定——哪怕一次拉扯失误,都会导致子细胞染色体数量异常,引发死亡或病变。但它的DNA序列却像被按下了快进键,进化速度是基因组平均水平的3倍。这就是「着丝粒悖论」的核心:功能越保守,序列越多变。
这次的研究终于给出了答案:不是着丝粒在瞎折腾,而是转座子的「入侵」逼着着丝粒蛋白一起进化。当Ty5转座子的序列被改造成着丝粒核心后,识别它的CBF3蛋白复合体也必须跟着调整自己的结构,才能精准抓住这个新的「挂扣」。这种DNA和蛋白的「协同进化」,让着丝粒在保持功能稳定的同时,DNA序列却能快速迭代。
我认为,这是整个研究最被低估的部分——它不仅解决了酵母的谜题,更给「着丝粒悖论」提供了普适性的解释:那些看似无用的基因组垃圾,其实是进化的「储备库」。当细胞的原有机制失效时,这些「垃圾」就能被紧急征用,变成新的功能结构,而蛋白则通过快速进化来适配这些新结构,最终达成新的平衡。
其实转座子的「逆袭」并非孤例。人类基因组里有一半以上的序列是转座子衍生的,其中一些已经被改造成了基因调控元件——比如我们熟悉的X染色体失活中心,核心序列就来自一种古老的转座子。甚至连大脑的发育,都有转座子在暗中参与:它们能在神经细胞里随机跳跃,制造神经元的遗传多样性,让每个人的大脑都独一无二。
但这次酵母的研究,第一次完整展现了转座子被驯化为核心染色体结构的全过程:从寄生的垃圾DNA,到过渡态的「原始点着丝粒」,再到极致精简的功能结构。更重要的是,它让我们看到了进化的「即兴发挥」——自然选择不会提前规划,它只会抓住每一个偶然的机会,把看似无用的东西,变成生命必需的零件。
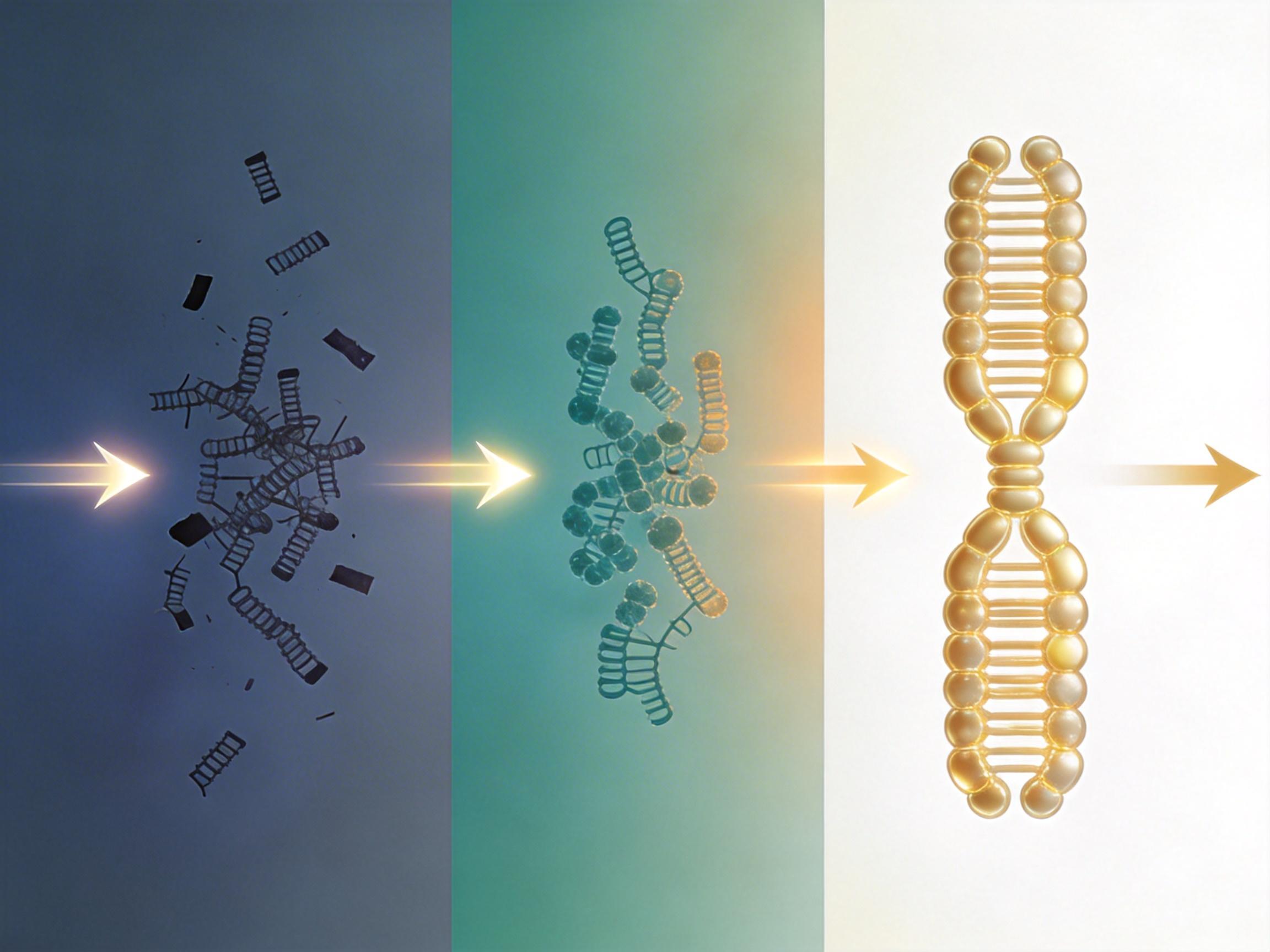
当然,这种「即兴发挥」也有风险。如果转座子跳错了位置,就可能导致基因突变或染色体异常,引发癌症等疾病。但正是这种风险与机遇并存的机制,让基因组能在稳定与创新之间找到平衡,推动生命不断进化。
当我们盯着显微镜下的酵母细胞,看着那些只有120个碱基对的着丝粒精准地勾住纺锤丝,把染色体拉向两个子细胞时,很难不感叹进化的神奇。那些曾被我们当成「垃圾」的DNA,其实是生命最隐秘的创新源泉。
垃圾DNA,是进化的隐藏手牌。
这个发现不仅破解了一个40年的科学谜题,更重新定义了我们对基因组的认知:基因组里没有真正的垃圾,只有尚未被发现价值的序列。未来,当我们再面对那些看似无用的DNA片段时,或许会多一份敬畏——谁知道它们会不会在几百万年后,变成生命的下一个核心结构?